Свойством рнк не является. Строение и функции рнк
Время, в которое мы живем, отмечено потрясающими переменами, огромным прогрессом, когда люди получают ответы на все новые и новые вопросы. Жизнь стремительно движется вперед, и то, что еще совсем недавно казалось невозможным, начинает претворяться в жизнь. Вполне возможно, что представляется сегодня сюжетом из жанра фантастики, скоро тоже приобретет черты реальности.
Одним из важнейших открытий во второй половине двадцатого столетия стали нуклеиновые кислоты РНК и ДНК, благодаря которым человек приблизился к разгадкам тайн природы.
Нуклеиновые кислоты
Нуклеиновые кислоты - это органические соединения, обладающие высокомолекулярными свойствами. В их состав входят водород, углерод, азот и фосфор.
Они были открыты в 1869 году Ф. Мишером, который исследовал гной. Однако тогда его открытию не придали особого значения. Лишь позже, когда эти кислоты обнаружили во всех животных и растительных клетках, пришло понимание огромной их роли.
Существуют два вида нуклеиновых кислот: РНК и ДНК (рибонуклеиновые и дезоксирибонуклеиновые кислоты). Настоящая статья посвящена рибонуклеиновой кислоте, но для общего понимания рассмотрим также, что собой представляет ДНК.
Что такое
ДНК — это состоящая из двух нитей, которые соединены по закону комплементарности водородными связями азотистых оснований. Длинные цепи закручены в спираль, один виток содержит почти десять нуклеотидов. Диаметр двойной спирали составляет два миллиметра, расстояние между нуклеотидами - около половины нанометра. Длина одной молекулы порой достигает нескольких сантиметров. Длина ДНК ядра человеческой клетки составляет почти два метра.
В структуре ДНК содержится вся ДНК обладает репликацией, что означает процесс, в ходе которого из одной молекулы образуются две совершенно одинаковые - дочерние.
Как уже было отмечено, цепь складывается из нуклеотидов, состоящих, в свою очередь, из азотистых оснований (аденина, гуанина, тимина и цитозина) и остатка кислоты фосфора. Все нуклеотиды различаются азотистыми основаниями. Водородная связь возникает не между всеми основаниями, аденин, к примеру, может соединяться только с тимином или гуанином. Таким образом, адениловых нуклеотидов в организме столько же, сколько тимидиловых, а число гуаниловых равно цитидиловым (правило Чаргаффа). Получается, что последовательность одной цепочки предопределяет последовательность другой, и цепи как бы зеркально отражают друг друга. Такая закономерность, где нуклеотиды двух цепей располагаются упорядоченно, а также соединяются избирательно, называется принципом комплементарности. Кроме водородных соединений, двойная спираль взаимодействует и гидрофобно.
Две цепи разнонаправлены, то есть расположены в противоположных направлениях. Поэтому напротив трех"-конца одной находится пяти"-конец другой цепи.
Внешне напоминает винтовую лестницу, перилом которой является сахарофосфатный остов, а ступеньками — комплементарные основания азота.
Что такое рибонуклеиновая кислота?

РНК — это нуклеиновая кислота с мономерами, называющимися рибонуклеотидами.
По химическим свойствам она очень похожа на ДНК, так как обе являются полимерами нуклеотидов, представляющих собой фосфолированный N-гликозид, который выстроен на остатке пентозы (пятиуглеродного сахара), с фосфатной группой пятого углеродного атома и основания азота при первом углеродном атоме.
Она представляет собой одну полинуклеотидную цепочку (кроме вирусов), которая намного короче, чем у ДНК.
Один мономер РНК — это остатки следующих веществ:
- основания азота;
- пятиуглеродного моносахарида;
- кислоты фосфора.
РНК имеют пиримидиновые (урацил и цитозин) и пуриновые (аденин, гуанин) основания. Рибоза является моносахаридом нуклеотида РНК.
Отличия РНК и ДНК

Нуклеиновые кислоты отличаются друг от друга следующими свойствами:
- количество ее в клетке зависит от физиологического состояния, возраста и органной принадлежности;
- ДНК содержит углевод дезоксирибозу, а РНК — рибозу;
- азотистое основание у ДНК — тимин, а у РНК — урацил;
- классы выполняют различные функции, но синтезируются на матрице ДНК;
- ДНК состоит из двойной спирали, а РНК — из одинарной цепи;
- для нее нехарактерны действующие у ДНК;
- в РНК больше минорных оснований;
- цепи существенно отличаются по длине.
История изучения
Клетка РНК впервые была открыта биохимиком из Германии Р. Альтманом при исследовании дрожжевых клеток. В середине двадцатого века была доказана роль ДНК в генетике. Лишь тогда описали и типы РНК, функции и так далее. До 80-90% массы в клетке приходится на р-РНК, образующих совместно с белками рибосому и участвующих в биосинтезе белка.
В шестидесятых годах прошлого столетия впервые предположили, что должен существовать некий вид, который несет в себе генетическую информацию для синтеза белка. После этого научно установили, что есть такие информационные рибонуклеиновые кислоты, представляющие комплементарные копии генов. Их еще называют матричными РНК.
В декодировании записанной в них информации участвуют так называемые транспортные кислоты.
Позже стали разрабатываться способы выявления последовательности нуклеотидов и устанавливаться структура РНК в пространстве кислоты. Так было обнаружено, что некоторые из них, которые назвали рибозимами, могут расщеплять полирибонуклеотидные цепи. Вследствие этого стали предполагать, что в то время, когда зарождалась жизнь на планете, РНК действовала и без ДНК и белков. При этом все превращения производились с ее участием.
Строение молекулы рибонуклеиновой кислоты
Почти все РНК - это одиночные цепи полинуклеотидов, которые, в свою очередь, состоят из монорибонуклеотидов — пуриновых и пиримидиновых оснований.
Нуклеотиды обозначают начальными буквами оснований:
- аденина (А), А;
- гуанина (G), Г;
- цитозина (С), Ц;
- урацила (U), У.
Они связаны между собой трех- и пятифосфодиэфирными связями.

Самое разное количество нуклеотидов (от нескольких десятков до десятков тысяч) входит в строение РНК. Они могут формировать вторичную структуру, состоящую в основном из коротких двуцепочных тяжей, которые образовались комплементарными основаниями.
Структура молекулы рибнуклеиновой кислоты
Как уже было сказано, у молекулы имеется однонитевое строение. РНК получает вторичную структуру и форму в результате взаимодействия нуклеотидов между собой. Это полимер, мономером которого является нуклеотид, состоящий из сахара, остатка кислоты фосфора и основания азота. Внешне молекула похожа на одну из цепей ДНК. Нуклеотиды аденин и гуанин, входящие в состав РНК, относятся к пуриновым. Цитозин и урацил являются пиримидиновыми основаниями.
Процесс синтеза
Чтобы молекула РНК синтезировалась, матрицей является молекула ДНК. Бывает, правда, и обратный процесс, когда новые молекулы дезоксирибонуклеиновой кислоты образуются на матрице рибонуклеиновой. Такое встречается при репликации некоторых видов вирусов.
Основой для биосинтеза могут служить также другие молекулы рибонуклеиновой кислоты. В ее транскрипции, которая происходит в ядре клетки, участвуют много ферментов, но самым значимым из них является РНК-полимераза.
Виды
В зависимости от вида РНК, функции ее также отличаются. Существуют несколько видов:
- информационная и-РНК;
- рибосомальная р-РНК;
- транспортная т-РНК;
- минорная;
- рибозимы;
- вирусные.

Информационная рибонуклеиновая кислота
Такие молекулы еще называют матричными. Они составляют в клетке примерно два процента от всего количества. В клетках эукариот они синтезируются в ядрах на ДНК-матрицах, переходя затем в цитоплазму и связываясь с рибосомами. Далее, они становятся матрицами для синтеза белка: к ним присоединяются транспортные РНК, которые несут аминокислоты. Так происходит процесс преобразования информации, которая реализуется в уникальной структуре белка. В некоторых вирусных РНК она к тому же является хромосомой.
Жакоб и Мано являются открывателями этого вида. Не имея жесткой структуры, ее цепь образует изогнутые петли. Не работая, и-РНК собирается в складки и сворачивается в клубок, а в рабочем состоянии разворачивается.
и-РНК несет в себе информацию о последовательности аминокислот в белке, который синтезируется. Каждая аминокислота закодирована в определенном месте при помощи генетических кодов, которым свойственны:
- триплетность — из четырех мононуклеотидов возможно выстроить шестьдесят четыре кодона (генетического кода);
- неперекрещиваемость — информация движется в одном направлении;
- непрерывность — принцип работы сводится к тому, что одна и-РНК — один белок;
- универсальность — тот или иной вид аминокислоты кодируется у всех живых организмов одинаково;
- вырожденность — известными являются двадцать аминокислот, а кодонов — шестьдесят один, то есть они кодируются несколькими генетическими кодами.
Рибосомальная рибонуклеиновая кислота
Такие молекулы составляют подавляющее большинство клеточных РНК, а именно от восьмидесяти до девяноста процентов от общего количества. Они соединяются с белками и формируют рибосомы — это органоиды, выполняющие синтез белков.
Рибосомы состоят на шестьдесят пять процентов из р-РНК и на тридцать пять процентов из белка. Эта полинуклеотидная цепь без труда изгибается вместе с белком.
Рибосома состоит из аминокислотного и пептидного участков. Они расположены на контактирующих поверхностях.
Рибосомы свободно передвигаются нужных местах. Они не очень специфичны и могут не только считывать информацию с и-РНК, но и образовывать с ними матрицу.
Транспортная рибонуклеиновая кислота
т-РНК наиболее изучены. Они составляют десять процентов клеточной рибонуклеиновой кислоты. Эти виды РНК связываются с аминокислотами благодаря специальному ферменту и доставляются на рибосомы. При этом аминокислоты переносятся транспортными молекулами. Однако бывает, что аминокислоту кодируют разные кодоны. Тогда переносить их будут несколько транспортных РНК.
Она сворачивается в клубочек, когда неактивна, а функционируя, имеет вид клеверного листа.
В ней различаются следующие участки:
- акцепторный стебель, имеющий последовательность нуклеотидов АЦЦ;
- участок, служащий для присоединения к рибосоме;
- антикодон, кодирующий аминокислоту, которая присоединена к этой т-РНК.
Минорный вид рибонуклеиновой кислоты
Недавно виды РНК пополнились новым классом, так называемыми малыми РНК. Они, скорее всего, являются универсальными регуляторами, которые включают или выключают гены в эмбриональном развитии, а также контролируют процессы внутри клеток.
Рибозимы также недавно выявлены, они активно принимают участие, когда кислота РНК ферментируется, являясь при этом катализатором.
Вирусные виды кислот
Вирус способен содержать либо рибонуклеиновую кислоту, либо дезоксирибонуклеиновую. Поэтому с соответствующими молекулами они называются РНК-содержащими. При попадании в клетку такого вируса происходит обратная транскрипция — на базе рибонуклеиновой кислоты появляются новые ДНК, которые встраиваются в клетки, обеспечивая существование и размножение вируса. В другом случае происходит образование комплиментарной на поступившей РНК. Вирусы белков, жизнедеятельность и размножение идет без ДНК, а лишь на основе информации, содержащейся в РНК вируса.
Репликация
В целях улучшения общего понимания необходимо рассмотреть процесс репликации, в результате которого появляются две идентичные молекулы нуклеиновой кислоты. Так начинается деление клетки.
В ней участвуют ДНК-полимеразы, ДНК-зависимые, РНК-полимеразы и ДНК-лигазы.
Процесс репликации состоит из следующих этапов:
- деспирализация — происходит последовательное раскручивание материнской ДНК, захватывающей всю молекулу;
- разрыв водородных связей, при котором цепи расходятся, и появляется репликативная вилка;
- подстройка дНТФ к освободившимся основаниям материнских цепей;
- отщепление пирофосфатов от дНТФ молекул и образование фосфорнодиэфирных связей за счет выделяющейся энергии;
- респирализация.
После образования дочерней молекулы делится ядро, цитоплазма и остальное. Таким образом, образуются две дочерние клетки, полностью получившие всю генетическую информацию.
Кроме этого, кодируется первичная структура белков, которые в клетке синтезируются. ДНК в этом процессе принимает косвенное участие, а не прямое, заключающееся в том, что именно на ДНК происходит синтез, участвующих в образовании белков, РНК. Этот процесс получил название транскрипции.
Транскрипция
Синтез всех молекул происходит во время транскрипции, то есть переписывании генетической информации с определенного оперона ДНК. Процесс в некоторых моментах похож на репликацию, а в других существенно отличается от нее.
Сходствами являются следующие части:
- начало идет с деспирализации ДНК;
- происходит разрыв водородных связей между основаниями цепей;
- к ним комплементарно подстраиваются НТФ;
- происходит образование водородных связей.
Отличия от репликации:
- при транскрипции расплетается лишь участок ДНК, соответствующий транскриптону, в то время как при репликации расплетению подвергается вся молекула;
- при транскрипции подстраивающиеся НТФ содержат рибозу, и вместо тимина урацил;
- информация списывается лишь с определенного участка;
- после образования молекулы водородные связи и синтезированная цепь разрываются, а цепь соскальзывает с ДНК.
Для нормального функционирования первичная структура РНК должна состоять только из списанных с экзонов ДНК-участков.
У только что образованных РНК начинается процесс созревания. Молчащие участки вырезаются, а информативные сшиваются, образуя полинуклеотидную цепь. Далее, каждый вид имеет присущие только ему превращения.
В и-РНК происходит присоединение к начальному концу. К конечному участку присоединяется полиаденилат.
В т-РНК модифицируются основания, образуя минорные виды.
У р-РНК также метилируются отдельные основания.
Защищают от разрушения и улучшают транспортировку в цитоплазму белки. РНК в зрелом состоянии с ними соединяются.
Значение дезоксирибонуклеиновых и рибонуклеиновых кислот

Нуклеиновые кислоты имеют огромное значение в жизнедеятельности организмов. В них хранится, переносится в цитоплазму и передается по наследству дочерним клеткам информация о белках, синтезирующихся в каждой клетке. Они присутствуют во всех живых организмах, стабильность этих кислот играет важнейшую роль для нормального функционирования как клеток, так и всего организма. Любые изменения в их строении приведут к клеточным изменениям.
Молекула РНК также полимер, мономерами которого является рибонуклеотиды, РНК представляет собой одноцепочную молекулу. Она построена таким же образом, как и одна из цепей ДНК. Нуклеотиды РНК похожи на нуклеотиды ДНК, хотя и не тождественны им. Их тоже четыре, и они состоят из осатков азотистого основания, пентозы и фосфорной кислоты. Три азотистых основания совершенно такие же, как в ДНК: А , Г и Ц . Однако вместо Т у ДНК в РНК присутствует близкое по строение пиримидиновое основание – урацил (У ). основное различие между ДНК и РНК – это характер углевода: в нуклотидах ДНК моносахарид – дезоксирибоза, а в РНК – рибоза. Связь между нуклеотидами осуществляется, как и в ДНК, через сахар и остаток фосфорной кислоты. В отличие от ДНК, содержание которой в клетках определенных организмов постоянно, содержание РНК в них колеблется. Оно заметно выше там, где происходит интенсивный синтез.
В отношении выполняемых функций различают несколько видов РНК.
Транспортная РНК (тРНК). Молекулы тРНк самые короткие: они состоят всего из 80-100 нуклеотидов. Молекулярная масса таких частиц равна 25-30 тыс. Транспортные РНК в основном содержатся в цитоплазме клетки. Функция их состоит в переносе аминокислот в рибосомы, к месту синтеза белка. Из общего содержания РНК клетк на долю тРНК приходится около 10%.
Рибосомная РНК (рРНК). Это крупные молекулы: в их состав входит 3-5 тыс. нуклеотидов, соотвественно их молекулярная масса достигает 1-1,5 млн. Рибосомные РНК составляют существенную часть рибосомы. Из общего содержания РНК в клетке на долю рРНК приходится около 90%.
Информационная РНК (иРНК), или матричная РНК (мРНК), содержится в ядре и цитоплазме. Функция ее состоит в переносе информации о структуре белка от ДНК к месту синтеза белка в рибосомах. На долю иРНК приходится примерно 0,5-1% от общего содержания РНК клетки. Размер иРНК колеблется в широких пределах – от 100 до 10000 нуклеотидов.
Все виды РНК синтезируются на ДНК, которая служитсвоего рода матрицей.
ДНК – носитель наследсвенной нформации.
Каждый белок представлен одной или несколькими полипиптидными цепями. Участок ДНК, несущий информацию об одной полипиптидной цепи, называют геном . Совокупность молекул ДНК клетки выполняет функцию носителя генетической информации. Генетическая информация передается как от материнской клетки дочерним клеткам, так и от родителей детям. Ген является единицей генетической , или наследственной, информации.
ДНК – носитель генетической информаци в клетке – непосредственного участия в синтезе белков не принимает. В клетках эукариот молекулы ДНК содержатся в хромосомах ядра и отделены ядерной оболочкой от цитоплазмы, где происходит синтез белков. К рибосомам – местам сборки белков – высылается из ядра несущий информацию посредник, способный пройти через поры ядерной оболочки. Таким посредником является информационная РНК (иРНК). По принципу комплементарности она синтезируется на ДНК при участие фермента, называемого РНК-полимеразой .

Информационная РНК – это однонитевая молекула, и транскрипция идет с одной цепи двунитевой молекулы ДНК. Она является копией не всей молекулы ДНК, а только части ее – одного гена у эукариот или группы рядом расположенных генов, несущих информацию о структуре белков, необходимых для выполнения одной функции, у прокариот. Такую группу генов называют опероном . В начале каждого оперона находится своего рода посадочная площадка для РНК-полимеразы, называемая промотором .это специфическая последовательность нуклеотидов ДНК, которую фермент «узнает» благодаря химическому сродству. Только присоединившись к промотору, РНК-полиммераза способна начать интез РНК. Доядя до конца оперона, фермент встречает сигнал (в виде определенной последоватльности нуклеотидов), означающий конец считывания. Готовая иРНК отходит от ДНК и направляется к месту синтеза белков.
В процессе транскрипции можно выделить четыре стадии: 1) связывание РНК -полимеразы с промотором; 2) инициация – начало синтеза. Оназаключается в образовании первой фосфодиэфирной связи между АТФ или ГТФ и вторым нуклеотидом синтезирующейся молекулы РНК; 3) элонгация – рост цепи РНК; т.е. последовательное присоединение нуклеотидов друг к другу в том порядке, в котором стоят комплементарные им нуклеотиды в транскрибируемой нити ДНК. Скорость элонгации 50 нуклеотидов в секунду; 4) терминация – завершение синтеза РНК.
Пройдя через поры ядерной оболочки, иРНК направляется к рибосомам, где осуществляется расшифровка генетической информации – перевод ее с «языка» нуклеотидов на «язык» аминокислот. Синтез полипептидных цепей по матрице иРНК, происходящий в рибосомах, называют трансляцией (лат. translation – перевод).
Аминокислоты, из котрых синтезируютсябелки, доставляются к рибосомам с помощью специальных РНК, называемых транспортными (тРНК). В клетке имеется столько же разных тРНК, сколько кодонов, шифрующих аминокислоты. На вершине «листа» каждой тРНК имеется последовательность трех нуклеотидов, комплементарных нуклеотидам кодона в иРНК. Ее называют антикодоном. Специальный фермент – кодаза – опознает тРНК и присоединяет к «черешку листа» аминокислоту – только ту, которая кодируется триплетом, комплементарным антикодону. На образование ковалентной связи между тРНК и «своей» аминокислотой затрачивается энергия одной молекулы АТФ.
 Для
того чтобы аминокислота включилась в
полипептидную цепь, она должна оторваться
от тРНК. Это становится возможным, когда
тРНК поступает на рибососму и антикодон
узнает свой кодон в иРНК. В рибосоме
имеется два участка для связывания двух
молекул тРНК. В один из этих участков,
называемый акцепторным
,
поступает тРНК с аминокислотой и
присоединяется к своему кодону (I).
Эта аминокислота присоединяет к себе
(акцептирует) растущую цепь белка (II)?
Между ними образуется пептидная связь.
тРНК, к которой теперь присоединяется
вместе с кодоном иРНК в донорный
участок рибосомы. В освободившийся
акцепторный участок приходит новая
тРНК, связанная с аминокислотой, которая
шифруется очередным кодоном (III).
Из донорного участка сюда вновь
переносится оторвавшаяся полипептидная
цепь и удлинняется еще на одно звено.
Аминокислоты в растущей цепи соединены
в той последовательности, в которой
расположены шифрующие их кодоны в иРНК.
Для
того чтобы аминокислота включилась в
полипептидную цепь, она должна оторваться
от тРНК. Это становится возможным, когда
тРНК поступает на рибососму и антикодон
узнает свой кодон в иРНК. В рибосоме
имеется два участка для связывания двух
молекул тРНК. В один из этих участков,
называемый акцепторным
,
поступает тРНК с аминокислотой и
присоединяется к своему кодону (I).
Эта аминокислота присоединяет к себе
(акцептирует) растущую цепь белка (II)?
Между ними образуется пептидная связь.
тРНК, к которой теперь присоединяется
вместе с кодоном иРНК в донорный
участок рибосомы. В освободившийся
акцепторный участок приходит новая
тРНК, связанная с аминокислотой, которая
шифруется очередным кодоном (III).
Из донорного участка сюда вновь
переносится оторвавшаяся полипептидная
цепь и удлинняется еще на одно звено.
Аминокислоты в растущей цепи соединены
в той последовательности, в которой
расположены шифрующие их кодоны в иРНК.
Когда на рибосоме оказывается один из трех триплетов (УАА, УАГ, УГА ), являющиеся «знаками препинания» между генами, ни одна тРНК не может занять место в акцепторном участке. Дело в том, что не существует антикодонов, комплементарных последовательностям нуклеотидов «знаков препинания». Оторвавшейся цепи не к чему присоединиться в акцепторном участке, и она покидает рибосому. Синтез белка завершен.
У прокариот синтез белков начинается с того, что кодон АУГ , расположенный на первом месте в копии с каждого гена, занимае в рибосоме такую позицию, что с ним взаимодействует антикодон особой тРНК, оединенной с формилментионином . Эта измененная форма аминокислоты метионина сразу попадает в донорный участок и выполняет роль заглавной буквы во фразе – с нее в бактериальной клетке начинается синтез любой полипептидной цепи. Когда триплет АУГ стоит не на первом месте, а внутри копии с гена, он кодирует аминокислоту метионин. После завершения синтеза полипептидной цепи формилметионин отщепляется от нее и в готовом белке отсуствует.
 Для
увеличения производства белков иРНК
часто проходит одновременно не по одной,
а по нескольким рибосомам. Акую структуру,
объединенную одной молекулой иРНК,
называют полисомой
.
На каждой рибосоме вэтом похожем на
нитку бус конвейере синтезируются
одинаковые белки.
Для
увеличения производства белков иРНК
часто проходит одновременно не по одной,
а по нескольким рибосомам. Акую структуру,
объединенную одной молекулой иРНК,
называют полисомой
.
На каждой рибосоме вэтом похожем на
нитку бус конвейере синтезируются
одинаковые белки.
Аминокислоты бесперебойно поставляются к рибосомам с помощью тРНК. Отдав аминокислоту, тРНК покидает рибосому и с помощью кодазы соединяется. Высокая слаженность всех «служб комбината» по производсву белов позволяет в течении нескольких секунд синтезировать полипептидные цепи, состоящие из сотен аминокислот.
Свойства генетического кода. Благодаря процессу транскрипции в клетке осуществляется передача информации от ДНК к белку
ДНК → иРНК → белок
Генетическая информация, содержащаяся в ДНК и в иРНК, заключена в последовательности расположения нуклеотидов в молекулах.
 Каким
же образом происходит перевод информации
с «языка» нуклеотидов на «язык»
аминокислот? Такой перевод осуществляется
с помощью генетического кода. Код,
или шифр
,
- это система символов для перевода
одной формы информации в другую.
Генетический
код
–это
система записи информации о
последовательности расположения
аминокислот в белках с помощью
последовательности расположения
нуклеотидов в иРНК.
Каким
же образом происходит перевод информации
с «языка» нуклеотидов на «язык»
аминокислот? Такой перевод осуществляется
с помощью генетического кода. Код,
или шифр
,
- это система символов для перевода
одной формы информации в другую.
Генетический
код
–это
система записи информации о
последовательности расположения
аминокислот в белках с помощью
последовательности расположения
нуклеотидов в иРНК.
Какими же свойствами обладает генетический код?
Код триплетен . В состав РНК входят четыре нуклеотида: А, Г, Ц, У. Если бы мы пытались обозначить одну аминокислоту одним нуклеотидом, то 16 из 20 аминокислот остались бы не зашифрованы. Двух буквенный код позволил бы зашифровать 16 аминокислот. Природа создала трехбуквенный, или триплетный, код. Это означает, что каждая из 20 аминокислот зашифрована последовательностью из трех нуклеотидов, называемой триплетом или кодоном.
Код вырожден. Это означает, что каждая аминокислота шифруется более чем одним кодоном. Исключения: метеонин и триптофан, каждая из которых кодируется одним триплетом.
Код однозначен. Каждый кодон шифрует только одну аминокислоту.
Между генами имеется «знаки препинания». В печатном тексте в конце каждой фразы стоит точка. Несколько связанных по смыслу фраз составляют абзац. На языке генетической информации таким абзацем являетсяоперон и комплементарная ему иРНК. Каждый ген в опероне прокариот или отдельный ген эукариот кодирует одну полипептидную цепочку – фразу. Так как в ряде случаев по матрице иРНК последовательно создается несколько разных полипептидных цепей, они должны быть отделены друг от друга. Для этого в генетическом годе имеются три специальных триплета – УАА, УАГ, УГА, каждый из которых обозначает прекращение синтеза одной полипептидной цепи. Таким образом, эти триплеты выполняют функцию знаков препинания. Они находятся в конце каждого гена.
Внутри гена нет «знаков препинания».
Код универсален. Генетический код един для всех живущих на Земле существ. У бактерий и грибов, пшеницы и хлопка, рыб и червей, лягушек и человека одни и те же триплеты кодируют одни и те же аминокислоты.

Принципы репликации ДНК. Преемственность генетического материала в поколениях клеток и организмов обеспечивается процессом репликации – удвоения молекул ДНК. Этот сложный процесс осуществляется комплексом нескольких ферментов и не обладающих каталитической активностью белов, необходимых для придания полинуклеотидным цепям нужной конформации. В результате репликации образуются две идентичные двойные спирали ДНК. Эти так называемые дочерние молекулы ничем не отличаются друг от друга и от исходной материнской молекулы ДНК. Репликация происходит в клетке перед делением, поэтому каждая дочерняя клетка получает точно такие же молекулы ДНК, какие имела материнская клетка. Процесс репликации основан на ряде принципов:
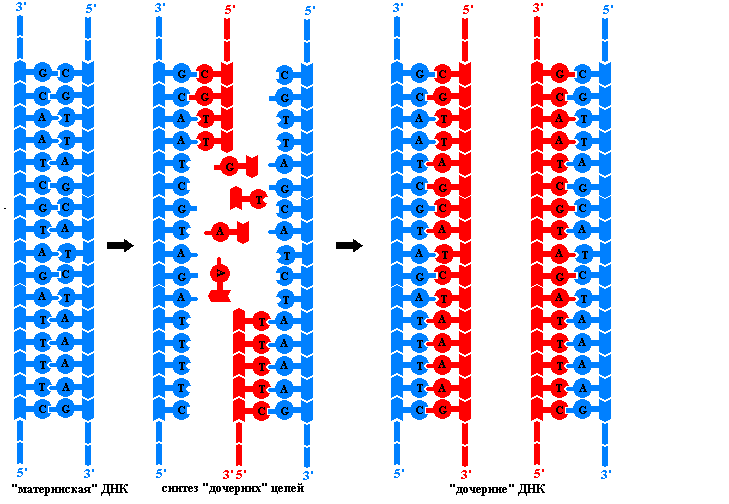
Только
в этом случае ДНК-полимеразы способна
двигаться по материнским нитям и
использовать их в качестве матриц для
безошибочного синтеза дочерних цепей.
Но полное раскручивание спиралей,
состоящих из многих миллионов пар
нуклеотидов, сопряжено со столь
значительным числом вращений и такими
энергетическими затратами, которые
невозможны в условиях клетки. Поэтому
репликация у эукариот начинается
одновременно в некоторых местах молекулы
ДНК. Участок между двумя точками, в
которых начинается синтез дочерних
цепей, называют репликоном
.
Он является единицей
репликации.


В каждой молекуле ДНК эукариотической клетки имеется много репликонов. В каждом репликоне можно видеть репликативную вилку – ту часть молекулы ДНК, которая под действием специальных ферментов уже расплелась. Каждая нить в вилке служит матрицей для синтеза комплементарной дочерней цепи. В ходе репликации вилка перемещается вдоль материнской молекулы, при этом расплетаются новые участки ДНК. Так как ДНК-полимеразы могут двигаться лишь в одном направлении вдоль матричных нитей, а нити ориентированы антипараллельно, то в каждой вилке одновременно ведут синтез два разных ферментативных комплекса. Причем в каждой вилке одна дочерняя (лидирующая) цепь растет непрерывно, а другая (отстающая) синтезируется отдельными фрагментами длинной в несколько нуклеотидов. Такие ферменты, названые в честь открывшего их японского ученого фрагментами Оказаки , сшиваются ДНК-лигазой, образуя непрерывную цепь. Механизм образования дочерних цепей ДНК фрагментами называют прерывистыми.
Потребность в затравке ДНК-полимераза не способна начинать синтез лидирующей цепи, ни синтез фрагментов Оказаки отстающей цепи. Она может лишь наращивать уже имеющуюся полинуклеотидную нить, последовательно присоединяя дезоксирибонуклеотиды к ее 3’-ОН концу. Откуда же берется начальный 5’-концевой участок растущей цепи ДНК? Его синтезирует на матрице ДНК особая РНК-полимераза, называемая праймазой (англ. Primer – затравка). Размер рибонуклеотидной затравки невелик (менее 20 нуклеотидов) в сравнении с размером цепи ДНК, образуемой ДНК-поимеразой. Выполнившая сво. Функци. РНК-затравка удаляется специальным ферментом, а образованная при эом брешь заделывается ДНК-полимеразой, использующей в качестве затравки 3’-ОН конец соседнего фрагмента Оказаки.
Проблема недорепликации концов линейных молекул ДНК. Удаление крайних РНК-праймеров, комплементрных 3’-концам обеих цепей линейной материнской молекулы ДНК, приводит к тому, что дочерние цепи оказываются короче 10-20 нуклеотидов. В этом и заключается проблема недорепликации концов линейных молекул.
Проблема недорепликации 3’-концов линейных молекул ДНК решается эукариотическими клетками с помощью специального фермента – теломеразы .
Теломераза является ДНК-полимеразой, достраивающей 3’-концылинейных молекул ДНК хромосом короткими повторяющимися последовательностями. Они, располагаясь друг за другом, образуют регулярную концевую структуру длинной до 10 тыс. нуклеотидов. Помимо белковой части, теломераза содержит РНК, выполняющую роль матрицы для наращивания ДНК повторами.
Схема удлинения концов молекул ДНК. Сначала происходит комплементарное связывание выступающего конца ДНК с матричным участком теломеразной РНК, затем теломераза наращивает ДНК, используя в качестве затравки ее 3’-ОН конец, а в качестве матрицы – РНК, входящую в состав фермента. Эта стадия называется элонгацией. После этого происходит транслокация, т.е. перемещение ДНК, удлиненной на один повтор, относительно фермента. Следом идет элонгация и очередная транслокация.
В результате образуются специализированные концевые структуры хромосом. Они состоят из многократно повторенных коротких последовательностей ДНК и специфических белков.
Относится к нуклеиновым кислотам. Молекулы-полимеры РНК намного меньше, чем у ДНК. Однако в зависимости от типа РНК количество входящих в них нуклеотидов-мономеров различается.
В состав нуклеотида РНК в качестве сахара входит рибоза, в качестве азотистого основания - аденит, гуанин, урацил, цитозин. Урацил по строению и химическим свойствам близок к тимину, который обычен для ДНК. В зрелых молекулах РНК многие азотистые основания модифицированы, поэтому в реальности разновидностей азотистых оснований в составе РНК намного больше.
Рибоза в отличие от дезоксирибозы имеет дополнительную -ОН-группу (гидроксильную). Это обстоятельство позволяет РНК легче вступать в химические реакции.
Главной функцией РНК в клетках живых организмов можно назвать реализацию генетической информации. Именно благодаря разным типам рибонуклеиновой кислоты генетический код считывается (транскрибируется) с ДНК, после чего на его основе синтезируются полипептиды (происходит трансляция). Итак, если ДНК в основном отвечает за хранение и передачу из поколения в поколение генетической информации (основной процесс – репликация), то РНК реализует эту информацию (процессы транскрипции и трансляции). При этом транскрипция происходит на ДНК, так что этот процесс относится к обоим типам нуклеиновых кислот и тогда с этой точки зрения можно сказать, что и ДНК отвечает за реализацию генетической информации.
При более подробном рассмотрении функции РНК намного разнообразнее. Ряд молекул РНК выполняют структурную, каталитическую и другие функции.
Существует так называемая гипотеза РНК-мира, согласно которой вначале в живой природе в качестве носителя генетической информации выступали только молекулы РНК, при этом другие молекулы РНК катализировали различные реакции. Данная гипотеза подтверждена рядом опытов, показывающих возможную эволюцию РНК. На это указывает и то, что ряд вирусов в качестве нуклеиновой кислоты, хранящей генетическую информацию, имеют молекулу РНК.
Согласно гипотезе РНК-мира ДНК появилась позже в процессе естественного отбора как более устойчивая молекула, что важно для хранения генетической информации.
Выделяют три основных типа РНК (кроме них есть и другие): матричная (она же информационная), рибосомальная и транспортная. Обозначаются они соответственно иРНК (или мРНК), рРНК, тРНК.
Информационная РНК (иРНК)
Почти все РНК синтезируются на ДНК в процессе транскрипции. Однако часто транскрипция упоминается как синтез именно информационной РНК (иРНК). Связано это с тем, что последовательность нуклеотидов иРНК в последствии определит последовательность аминокислот синтезируемого в процессе трансляции белка.
Перед транскрипцией нити ДНК расплетаются, и на одной из них с помощью комплекса белков-ферментов синтезируется РНК по принципу комплементарности, так же как это происходит при репликации ДНК. Только напротив аденина ДНК к молекуле РНК присоединяется нуклеотид, содержащий урацил, а не тимин.
На самом деле на ДНК синтезируется не готовая информационная РНК, а ее предшественник - пре-иРНК. Предшественник содержит участки последовательности нуклеотидов, которые не кодируют белок и которые после синтеза пре-иРНК вырезаются при участии малых ядерных и ядрышковых РНК («дополнительные» типы РНК). Эти удаляющиеся участки называются интронами . Остающиеся части иРНК называются экзонами . После удаления интронов экзоны сшиваются между собой. Процесс удаления интронов и сшивания экзонов называется сплайсингом . Усложняющей жизнь особенностью является то, что можно вырезать интроны по-разному, в результате получатся разные готовые иРНК, которые будут служить матрицами для разных белков. Таким образом, вроде бы один ген ДНК может играть роль нескольких генов.
Следует отметить, что у прокариотических организмов сплайсинга не происходит. Обычно их иРНК сразу после синтеза на ДНК готова к трансляции. Бывает, что пока конец молекулы иРНК еще транскрибируется, на ее начале уже сидят рибосомы, синтезирующие белок.
После того как пре-иРНК созревает в информационную РНК и оказывается вне ядра, она становится матрицей для синтеза полипептида. При этом на нее «насаживаются» рибосомы (не сразу, какая-то оказывается первой, другая - второй и т. д.). Каждая синтезирует свою копию белка, т. е. на одной молекуле РНК могут синтезироваться сразу несколько одинаковых белковых молекул (понятно, что каждая будет находиться на своей стадии синтеза).
Рибосома, передвигаясь от начала иРНК к ее концу, считывает по три нуклеотида (хотя вмещает шесть, т. е. два кодона) и присоединяет соответствующую транспортную РНК (имеющую соответствующий кодону антикодон), к которой присоединена соответствующая аминокислота. После этого с помощью активного центра рибосомы ранее синтезированная часть полипептида, соединенная с предшествующей тРНК, как-бы «пересаживается» (образуется пептидная связь) на аминокислоту, прикрепленную к только что пришедшей тРНК. Таким образом, молекула белка постепенно увеличивается.
Когда молекула информационной РНК становится не нужна, клетка ее разрушает.
Транспортная РНК (тРНК)
Транспортная РНК - это достаточно маленькая (по меркам полимеров) молекула (количество нуклеотидов бывает разным, в среднем около 80-ти), во вторичной структуре имеет форму клеверного листа, в третичной сворачивается в нечто подобное букве Г.

Функция тРНК - присоединение к себе соответствующей своему антикодону аминокислоты. В дальнейшем соединение с рибосомой, находящейся на соответствующем антикодону кодоне иРНК, и «передача» этой аминокислоты. Обобщая, можно сказать, что транспортная РНК переносит (на то она и транспортная) аминокислоты к месту синтеза белка.
Живая природа на Земле использует всего около 20-ти аминокислот для синтеза различных белковых молекул (на самом деле аминокислот куда больше). Но поскольку, согласно генетическому коду, кодонов больше 60-ти, то каждой аминокислоте может соответствовать несколько кодонов (на самом деле какой-то больше, какой-то меньше). Таким образом, разновидностей тРНК больше 20, при этом разные транспортные РНК переносят одинаковые аминокислоты. (Но и тут не так все просто.)
Рибосомная РНК (рРНК)
Рибосомную РНК часто также называют рибосомальной РНК. Это одно и то же.
Рибосомная РНК составляет около 80% всей РНК клетки, так как входит в состав рибосом, коих в клетке бывает достаточно много.
В рибосомах рРНК образует комплексы с белками, выполняет структурную и каталитическую функции.
В состав рибосомы входят несколько разных молекул рРНК, отличающиеся между собой как по длине цепи, вторичной и третичной структуре, выполняемым функциям. Однако их суммарная функция - это реализация процесса трансляции. При этом молекулы рРНК считывают информацию с иРНК и катализируют образование пептидной связи между аминокислотами.
Нуклеиновые кислоты - высокомолекулярные вещества, состоящие из мононуклеотидов, которые соединены друг с другом в полимерную цепочку с помощью 3",5"- фосфодиэфирных связей и упакованы в клетках определенным образом.
Нуклеиновые кислоты - биополимеры двух разновидностей: рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК) . Каждый биополимер состоит из нуклеотидов, различающихся по углеводному остатку (рибозе, дезоксирибозе) и одному из азотистых оснований (урацил, тимин). Соответственно этим различиям нуклеиновые кислоты и получили свое название.
Структура рибонуклеиновой кислоты
Первичная структура РНК
Молекула РНК представляют собой линейные (т. е. неразветвленные) полинуклеотиды с аналогичным ДНК принципом организации. Мономерами РНК являются нуклеотиды, состоящие из фосфорной кислоты, углевода (рибозы) и азотистого основания, соединенные 3", 5"-фосфодиэфирными связями. Полинуклеотидные цепи молекулы РНК полярны, т.е. имеют различимые 5’- и 3"-концы. При этом, в отличие от ДНК, РНК является одноцепочечной молекулой. Причиной такого отличия служат три особенности первичной структуры:- РНК в отличие от ДНК содержит вместо дезоксирибозы рибозу, которая имеет дополнительную гидроксигруппу. Гидроксигруппа делает двухцепочечную структуру менее компактной
- Среди четырех главных, или мажорных, азотистых оснований (А, Г, Ц и У) вместо тимина содержится урацлл, отличающийся от тимина лишь отсутствием метильной группы в 5-м положении. Благодаря этому уменьшается сила гидрофобного взаимодействия в комплементарной паре A-У, что тоже снижает вероятность образования устойчивых двухцепочечных молекул.
- Наконец, в РНК (особенно в тРНК) высоко содержание т. н. минорных оснований и нуклеозидов. Среди них дигидроуридин (в урациле нет одной двойной связи), псевдоуридин (урацил иначе, чем обычно, связан с рибозой), диметиладенин и диметилгуанин (в азотистых основаниях по две дополнительных метильных группы) и многие другие. Почти все эти основания не могут участвовать в комплементарных взаимодействиях. Так, метильные группы в диметиладенине (в отличие от тимина и 5-метилцитозина) находятся при таком атоме, который в паре A-У образует водородную связь; следовательно, теперь данная связь замкнуться не может. Это тоже препятствует образованию двухцепочечных молекул.
Таким образом, широко известные отличия состава РНК от ДНК имеют огромное биологическое значение: ведь свою функцию молекулы РНК способны выполнять только в одноцепочечном состоянии, что наиболее очевидно для мРНК: трудно представить, как бы могла двухцепочечная молекула транслироваться на рибосомах.
Вместе с тем, оставаясь одиночной, в некоторых участках цепь РНК может образовывать петли, выступы или "шпильки", с двухцепочечной структурой (рис.1.). Эта структура стабилизирована взаимодействием оснований в парах А::У и Г:::Ц. Однако могут образовываться и "не правильные" пары (например, Г У), а в некоторых местах "шпильки" и вообще не происходит никакого взаимодействия. В составе таких петель может содержаться (особенно в тРНК и рРНК) до 50 % всех нуклеотидов. Общее же содержание нуклеотидов в РНК варьирует от 75 единиц до многих тысяч. Но даже самые крупные РНК на несколько порядков короче хромосомных ДНК.
Первичная структура мРНК скопирована с участка ДНК, содержащего информацию о первичной структуре полипептидной цепи. Первичная структура остальных типов РНК (тРНК, рРНК, редкие РНК) является окончательной копией генетической программы соответствующих генов ДНК.
Вторичная и третичная структуры РНК
Рибонуклеиновые кислоты (РНК) - однонитевые молекулы, поэтому в отличие от ДНК их вторичная и третичная структуры нерегулярны. Эти структуры, определяемые как пространственная конформация полинуклеотидной цепи, формируются в основном за счет водородных связей и гидрофобных взаимодействий между азотистыми основаниями. Если для молекулы нативной ДНК характерна устойчивая спираль, то структура РНК более многообразна и лабильна. Рентгеноструктурный анализ показал, что отдельные участки полинуклеотидной цепи РНК, перегибаясь, навиваются сами на себя с образованием внутриспиральных структур. Стабилизация структур достигается за счет комплементарных спариваний азотистых оснований антипараллельных участков цепи; специфическими парами здесь являются А-U, G-С и, реже, G–U. Благодаря этому в молекуле РНК возникают как короткие, так и протяженные биспиральные участки, принадлежащие одной цепи; эти участки носят название шпилек. Модель вторичной структуры РНК со шпилькообразными элементами была создана в конце 50-х - начале 60-х гг. XX в. в лабораториях А. С. Спирина (Россия) и П. Доти (США).
| Некоторые виды РНК | ||
| Виды РНК | Размер в нуклеотидах | Функция |
| gРНК - геномные РНК | 10000-100000 | |
| mРНК - информационные (матричные) РНК | 100-100000 | передает информацию о структуре белка с молекулы ДНК |
| tPHK - транспортные РНК | 70-90 | транспортирует аминокислоты к месту синтеза белка |
| rРНК - рибосомные РНК | несколько дискретных классов от 100 до 500000 | содержится в рибосомах, участвует в поддержании структуры рибосомы |
| sn-PHK - малая ядерная РНК | 100 | удаляет интроны и ферментативно соединяет экзоны в мРНК |
| sno-РНК - малая ядрышковая РНК | участвует в направлении или проведении модификаций оснований в рРНК и малой ядерной РНК, таких, как, например, метилирование и псевдоуридинизация. Большинство малых ядрышковых РНК находятся в интронах других генов | |
| srp-РНК - сигналраспознающая РНК | распознаёт сигнальную последовательность белков, предназначенных для экспрессии, и участвует в их переносе через цитоплазматическую мембрану | |
| mi-РНК - микро-РНК | 22 | контролируют трансляцию структурных генов путём комплементарного связывания с 3"-концами нетранслируемых участков иРНК |
Образование спиральных структур сопровождается гипохромным эффектом - уменьшением оптической плотности образцов РНК при 260 нм. Разрушение этих структур происходит при понижении ионной силы раствора РНК или при его нагревании до 60-70 °С; оно также называется плавлением и объясняется структурным переходом спираль - хаотический клубок, что сопровождается увеличением оптической плотности раствора нуклеиновой кислоты.
В клетках существуют несколько типов РНК:
- информационная (или матричная) РНК (иРНК или мРНК) и её предшественница - гетерогенная ядерная РНК (г-я-РНК)
- транспортная РНК (т-РНК) и ее предшественница
- рибосомная (р-РНК) и ее предшественница
- малая ядерная РНК (sn-PHK)
- малая ядрышковая РНК (sno-PHK)
- сигналраспознающая РНК (srp-PHK)
- микро-РНК (mi-PHK)
- митохондриальная РНК (т+ РНК).

Гетерогенная ядерная и информационная (матричная) РНК
Гетерогенная ядерная РНК свойственна исключительно эукариотам. Она является предшественницей информационной РНК (и-РНК), которая переносит генетическую информацию от ядерной ДНК к цитоплазме. Гетерогенная ядерная РНК (пре-мРНК) открыта советским биохимиком Г. П. Георгиевым. Количество видов г-я-РНК равно количеству генов, так как она служит прямой копией кодирующих последовательностей генома, в силу чего имеет копии палиндромов ДНК, поэтому ее вторичная структура содержит шпильки и линейные участки. В процессе транскрипции РНК с ДНК ключевую роль играет фермент РНК-полимераза II.
Информационная РНК образуется в результате процессинга (созревания) г-я-РНК, при котором происходят отсечение шпилек, вырезание некодирующих участков (интронов) и склеивание кодирующих экзонов.
Информационная РНК (и-РНК) представляет собой копию определенного участка ДНК и выполняет роль переносчика генетической информации от ДНК к месту синтеза белка (рибосомы) и непосредственно участвует в сборке его молекул.
Зрелая матричная РНК имеет несколько областей с различной функциональной ролью (рис.)
- на 5"-конце находится т.н. "колпачок" или кэп - участок из одного-четырех модифицированных нуклеотидов. Такая структура защищает 5"-конец м-РНК от эндонуклеаз
- за "колпачком" идет 5"-нетранслируемая область - последовательность из нескольких десятков нуклеотидов. Она комплементрана одному из отделов той р-РНК, которая входит в малую субъединицу рибосомы. За счет этого она служит для первичного связывания м-РНК с рибосомой, но сама не транслируется
- инициирующий кодон - АУГ, кодирующий метионин. Во всех м-РНК инициирующий кодон одинаков. С него начинается трансляция (считывание) м-РНК. Если после синтеза пептидной цепи метионин не нужен, то он, как правило, отщепляется с ее N-конца.
- За инициирующим кодоном следует кодирующая часть, которая содержит информацию о последовательности аминокислот в белке. У эукариот зрелые м-РНК являются
моноцистронными, т.е. каждая из них несет информацию о структуре только одной полипептидной цепи.
Другое дело, что иногда пептидная цепь вскоре после образования на рибосоме разрезается на несколько более мелких цепей. Так бывает, например, при синтезе инсулина и целого ряда олигопептидных гормонов.
Кодирующая часть зрелой м-РНК эукариот лишена интронов - каких-либо вставочных некодирующих последовательностей. Иными словами, имеется непрерывная последовательность смысловых кодонов, которая должна читаться в направлении 5" ->3".
- По окончании этой последовательности находится кодон терминации - один из трех "бессмысленных" кодонов: УАА, УАГ или УГА (см. табл. генетического кода ниже).
- За этим кодоном может следовать еще 3"-нетранслируемый участок, значительно превышающий по длине 5’-нетранслируемую область.
- Наконец, почти все зрелые мРНК эукариот (кроме гистоновых мРНК) на 3"-конце содержат поли(А)-фрагмент из 150-200 адениловых нуклеотидов
3"-нетранслируемый участок и поли(А)-фрагмент имеют отношение к регуляции продолжительности жизни м-РНК, поскольку разрушение м-РНК осуществляется 3"-экзонуклеазами. После окончания трансляции м-РНК от поли(А)-фрагмента отщепляются 10-15 нуклеотидов. Когда данный фрагмент исчерпывается, начинает разрушаться значащая часть мРНК (если отсутствует 3"-нетранслируемый участок).
Общее количество нуклеотидов в мРНК обычно варьирует в пределах нескольких тысяч. При этом на кодирующую часть иногда может приходиться лишь 60-70 % нуклеотидов.
В клетках молекулы мРНК практически всегда связаны с белками. Последние, вероятно, стабилизируют линейную структуру мРНК, т. е. предупреждают образование в кодирующей части "шпилек". Кроме того, белки могут защищать м-РНК от преждевременного разрушения. Такие комплексы мРНК с белками иногда называют информосомами.
Транспортная РНК в цитоплазме клетки переносит аминокислоты в активированной форме к рибосомам, где они соединяются в пептидные цепи в определенной последовательности, которую задает РНК-матрица (мРНК). В настоящее время известны данные о нуклеотидной последовательности более чем 1700 видов тРНК из прокариотических и эукариотических организмов. Все они имеют общие черты как в их первичной структуре, так и в способе складывания полинуклеотидной цепи во вторичную структуру за счет комплементарного взаимодействия входящих в их структуру нуклеотидов.

Транспортная РНК в своем составе содержит не больше 100 нуклеотидов, среди которых отмечается высокое содержание минорных, или модифицированных, нуклеотидов. Первой полностью расшифрованной транспортной РНК была аланиновая РНК, выделенная из дрожжей. Анализ показал, что аланиновая РНК состоит из 77 нуклеотидов, расположенных в строго определенной последовательности; в их состав входят так называемые минорные нуклеотиды, представленные нетипичными нуклеозидами
Аланиновая тРНК содержит 9 необычных оснований с одной или несколькими метильными группами, которые присоединяются к ним ферментативным путем уже после образования фосфодиэфирных связей между нуклеотидами. Эти основания неспособны к образованию обычных пар; возможно, они служат для того, чтобы препятствовать спариванию оснований в определенных частях молекулы и таким образом обнажать специфические химические группы, которые образуют вторичные связи с информационной РНК, рибосомой или, быть может, с ферментом, необходимым для присоединения определенной аминокислоты к соответствующей транспортной РНК. Известная последовательность нуклеотидов в тРНК по существу означает, что известна также его последовательность в генах, на которых эта тРНК синтезируется. Эту последовательность можно вывести основываясь на правилах специфического спаривания оснований, установленных Уотсоном и Криком. В 1970 году была синтезирована полная двухцепочечная молекула ДНК с соответсвующей последовательностью из 77 нуклеотидов, и оказалось, что она может служить матрицей для построения аланиновой транспортной РНК. Это был первый искусственно синтезированный ген. |

Транскрипция тРНК
Транскрипция молекул т-РНК происходит с кодирующих её последовательностей в ДНК при участии фермента РНК-полимеразы III. В ходе транскрипции формируется первичная структура тРНК в виде линейной молекулы. Формирование начинается с составления РНК-полимеразой последовательности нуклеотидов в соответствии с геном, содержащим информацию о данной транспортной РНК. Эта последовательность представляет из себя линейную полинуклеотидную цепь, в которой нуклеотиды следуют друг за другом. Линейная полинуклеотидная цепь является первичной РНК, предшественницей тРНК, включающей в себя интроны - неинформативные излишки нуклеотидов. На этом уровне организации пре-тРНК не функциональна. Образуясь в разных местах ДНК хромосом пре-тРНК содержит излишки примерно в 40 нуклеотидов по сравнению со зрелой тРНК.
Вторым этапом вновь синтезированная предшественница тРНК проходит послетранскрипционное созревание или процессинг. В ходе процессинга удаляются неинформативные излишки в пре-РНК и образуются зрелые, функциональные молекулы РНК.
Процессинг пре-тРНК
Процессинг начинается с того, что в транскрибте образуется внутримолекулярные водородные связи и молекула тРНК принимает форму клеверного листа. Это вторичный уровень организации тРНК, на котором молекула тРНК еще не функциональна. Далее происходит вырезание неинформативных участков из пре-РНК, сращивание информативных участков "разорванных генов" - сплайсинг и модификация 5"- и 3"-концевых участков РНК.
Вырезание неинформативных участков пре-РНК осуществляется с помощью рибонуклеаз (экзо- и эндонуклеаз). После удаления излишков нуклеотидов происходит метилирование оснований тРНК. Реакция осуществляется метилтрансферазами. В роли донора метильных групп выступает S-аденозилметионин. Метилирование препятствует разрушению тРНК нуклеазами. Окончательно зрелая тРНК образуется путем присоединения специфической тройки нуклеотидов (акцепторного конца) - ЦЦА, которое осуществляется специальной РНК-полимеразой.
По завершении процессинга во вторичной структуре вновь образуются дополнительные водородные связи за счет которых тРНК переходит на третичный уровень организации и принимает вид так называемой L-формы. В таком виде тРНК уходит в гиалоплазму.
Строение тРНК
В основе структуры транспортной РНК лежит цепочка нуклеотидов. Однако в силу того, что любая цепочка нуклеотидов имеет положительно и отрицательно заряженные части, она не может находиться в клетке в развернутом состоянии. Эти заряженные части притягиваясь друг к другу легко образуют между собой водородные связи по принципу комплементарности. Водородные связи причудливо скручивают нить т-РНК и удерживают ее в таком положении. В результате этого вторичная структура т-РНК имеет вид "клеверного листа" (рис.), содержащего в своей структуре 4 двухцепочечных участка. Высокое содержание минорных или модифицированных нуклеотидов, отмечаемых в цепи тРНК и неспособных к комплементарным взаимодействиям, формирует 5 одноцепочечных участков.
Т.о. вторичная структура т-РНК образуется вследствие внутрицепочечного спаривания комплементарных нуклеотидов отдельных участков тРНК. Участки тРНК, не вовлекаемые в образование водородных связей между нуклеотидами, образуют петли или линейные звенья. В тРНК выделяют следующие структурные участки:
- Акцепторный участок (конец) , состоящий из четырех линейно расположенных нуклеотидов, три из которых имеют во всех типах тРНК одинаковую последовательность - ЦЦА. Гидроксил 3"-ОН аденозина свободен. К нему присоединяется карбоксильной группой аминокислота, отсюда и название этого участка тРНК - акцепторный. Связанную с 3"-гидроксильной группой аденозина аминокислоту тРНК доставляет к рибосомам, где происходит синтез белка.
- Антикодоновая петля , обычно образуемая семью нуклеотидами. Она содержит специфический для каждой тРНК триплет нуклеотидов, называемый антикодоном. Антикодон тРНК по принципу комплементарности спаривается с кодоном мРНК. Кодон-антикодоновое взаимодействие определяет порядок расположения аминокислот в полипептидной цепи во время сборки ее в рибосомах.
- Псевдоуридиловая петля (или ТΨС-петля) , состоящая из семи нуклеотидов и обязательно содержащая остаток псевдоуридиловой кислоты. Предполагают, что псевдоуридиловая петля участвует в связывании тРНК с рибосомой.
- Дигидроуридиновая, или D-петля , состоящая обычно из 8-12 нуклеотидных остатков, среди которых обязательно имеется несколько остатков дигидроуридина. Считают, что D-петля необходима для связывания с аминоацил-тРНК-синтетазой, которая участвует в узнавании аминокислотой своей тРНК (см. "Биосинтез белка"),
- Добавочная петля , которая варьирует по размерам и составу нуклеотидов у разных тРНК.
Третичная структура т-РНК уже не имеет формы клеверного листа. За счет образования водородных связей между нуклеотидами из разных частей "листа клевера" его лепестки заворачиваются на тело молекулы и удерживаются в таком положении дополнительно ван-дер-ваальсовыми связями, напоминая собой форму буквы Г или L. Наличие стабильной третичной структуры является еще одной особенностью т-РНК, в отличие от длинных линейных полинуклеотидов м-РНК. Понять, как именно изгибаются различные части вторичной структуры т-РНК при образовании третичной структуры можно по рис., сопоставив цвета схемы вторичной и третичной структуры т-РНК.


Транспортные РНК (т-РНК) переносят аминокислоты из цитоплазмы в рибосомы в процессе синтеза белка. Из таблицы с генетическим кодом видно, что каждая аминокислота кодируется несколькими последовательностями нуклеотидов, поэтому каждой аминокислоте соответствуют свои транспортные РНК. В результате этого существует большое разнообразие т-РНК: от одного до шести видов для каждой из 20 аминокислот. Виды тРНК, способные связывать одну и ту же аминокислоту, называются изоакцепторными (например аланин может быть присоединен к т-РНК, антикодон которой будет комплементарен кодонам GCU, GCC, GCA, GCG). Специфичность тРНК обозначается верхним индексом, например: тРНК Ala .
Для процесса синтеза белка главными функциональными частями т-РНК являются: антикодон - последовательность нуклеотидов, находящихся на антикодоновой петле, комплементарных кодону информационной РНК (и-РНК) и акцепторная часть - противоположный антикодону конец т-РНК, к которому присоединяется аминокислота. Последовательность оснований в антикодоне напрямую зависит от вида аминокислоты, прикреплённой к 3"-концу. Так, например, т-РНК, антикодон которой имеет последовательность 5"-ЦЦА-3", может нести только аминокислоту триптофан. Следует отметить, что данная зависимость лежит в основе передачи генетической информации, носителем которой выступает т-РНК.
В процессе синтеза белка т-РНК антикодоном распознает трехбуквенную последовательность генетического кода (кодона) и-РНК, сопоставляя ей единственную соответствующую аминокислоту, закрепленную на другом конце тРНК. Только в случае комплементарности антикодона к участку мРНК транспортная РНК может к ней присоединиться и отдать переносимую аминокислоту на формирование протеиновой цепочки. Взаимодействие т-РНК и и-РНК происходит в рибосоме, которая также является активным участником трансляции.
Распознавание т-РНК своей аминокислоты и кодона и-РНК происходит определенным образом:
- Связывание "своей" аминокислоты с т-РНК происходит с помощью фермента - специфической аминоацил-тРНК-синтетазы
Существуют большое разнообразие аминоацил-тРНК-синтетаз - по числу тРНК, используемых аминокислотами. Сокращенно их называют АРСазы. Аминоацил-тРНК-синтетазы крупные молекулы (мол.масса 100 000 - 240 000), имеющие четвертичную структуру. Они специфически узнают тРНК и аминокислоты и катализируют их соединение. Для этого процесса требуется АТФ, энергия которой используется на активирование аминокислоты с карбоксильного конца и присоединение ее к гидроксилу (3"-ОН) аденозина акцепторного конца (ЦЦА) тРНК. Считается, что в молекуле каждой аминоацил-тРНК-синтетазы имеются центры связывания по крайней мере три центра связывания: для аминокислоты, изоакцепторных тРНК и АТФ. В центрах связывания происходит образование ковалентной связи, при соответствии аминокислоты тРНК, и гидролиз такой связи в случае их несоответствия (присоединения к тРНК "не той" аминокислоты).
АРСазы обладают способностью выборочно использовать при узнавании ассортимент тРНК для каждой аминокислоты, т.е. ведущим звеном узнавания является аминокислота, а к ней подгоняется своя тРНК. Далее тРНК путем простой диффузии переносит присоединенную к ней аминокислоту к рибосомам, где происходит сборка белка из аминокислот, поступающих в виде разных аминоацил-тРНК.

Связывание аминокислоты с тРНК
Связывание тРНК и аминокислоты происходит следующим образом (рис.): к аминоацил-тРНК-синтетазе присоединяется аминокислота и молекула АТФ. Для последующего аминоацелирования молекула АТФ высвобождает энергию, отщепляя две фосфатные группы. Оставшийся АМФ (аденозинмонофосфат) присоединяется к аминокислоте, подготавливая ее к соединению с акцепторным участком тРНК - акцепторной шпилькой. После чего синтетаза присоединяет к себе соответствующую аминокислоте родственную тРНК. На этом этапе происходит проверка соответствия тРНК синтетазе. В случае сооответствия тРНК плотно присоединяется к синтетазе, изменяя ее структуру, что приводит к запуску процесса аминоацелирования - присоединения аминокислоты к тРНК.
Аминоацилирование происходит в процессе замены присоединенной к аминокислоте молекулы АМФ на молекулу тРНК. После этой замены АМФ покидает синтетазу, а тРНК задерживается для последней проверки аминокислоты.
Проверка соответствия тРНК присоединенной аминокислоте
Модель синтетазы для проверки соответствия тРНК присоединенной аминокислоте предполагает наличие двух активных центров: синтетического и коррекционного. В синтетическом центре происходит присоединение тРНК к аминокислоте. Акцепторный участок тРНК, захваченной синтетазой, вначале контактирует с синтетическим центром, в котором уже размещена аминокислота, соединенная с АМФ. Этот контакт акцепторного участка тРНК придает ему неестественный изгиб до момента присоединения аминокислоты. После того, как происходит присоединение аминокислоты с акцепторному участку тРНК, необходимость нахождения данного участка в синтетическом центре отпадает, тРНК распрямляется и перемещает присоединенную к ней аминокислоту в коррекционный центр. При несовпадении размеров молекулы аминокислоты, присоединенной к тРНК, и размеров коррекционного центра, аминокислота опознается как неправильная и отсоединяется от тРНК. Синтетаза готова к следующему циклу. При совпадении размеров молекулы аминокислоты, присоединенной к тРНК, и размеров коррекционного центра, заряженная аминокислотой тРНК освобождается: она готова сыграть свою роль в трансляции протеина. А синтетаза готова присоединить новые аминокислоту и тРНК, и начать повторный цикл.
Соединение несоответствующей аминокислоты с синтетазой в среднем происходит в 1-м случае из 50 тыс., а с ошибочной тРНК всего лишь один раз на 100 тысяч присоединений.
- Взаимодействие кодона м-РНК и антикодона т-РНК происходит по принципу комплементарности и антипараллельности
Взаимодействие тРНК с кодоном мРНК по принципу комплементраности и антипараллельности означает: поскольку смысл кодона мРНК читается в направлении 5"->3", то антикодон в тРНК должен читаться в направлении 3"->5". При этом первые два основания кодона и антикодона спариваются строго комплементарно, т. е. образуются только пары А У и Г Ц. Спаривание же третьих оснований может отступать от этого принципа. Допустимые пары определяются схемой:
Из схемы вытекает следующее.
- Молекула тРНК связывается только с 1-м типом кодона, если третий нуклеотид в ее антикодоне - Ц или А
- тРНК связывается с 2-мя типами кодонов, если антикодон заканчивается на У или Г.
- И, наконец, тРНК связывается с 3-мя типами кодонов, если антикодон кончается на И (инозиновый нуклеотид); такая ситуация, в частности, в аланиновой
тРНК.
Отсюда, в свою очередь, следует, что для узнавания 61 смыслового кодона требуется, в принципе, не такое же, а меньшее количество разных тРНК.
Рибосомальная РНК

Рибосомальные РНК являются основой для формирования субъединиц рибосом. Рибосомы обеспечивают пространственное взаиморасположение мРНК и тРНК в процессе синтеза белка.
Каждая рибосома состоит из большой и малой субъединиц. Субъединицы включают в себя большое количество белков и рибосомальные РНК, которые не подвергаются трансляции. Рибосомы, как и рибосомальные РНК, различаются по коэффициенту седиментации (осаждения), измеряемому в единицах Сведберга (S). Данный коэффициент зависит от скорости осаждения субъединиц при центрифугировании в насыщенной водной среде.
Каждая рибосома эукариот имеет коэффициент седиментации, равный 80S, и ее принято обозначать как 80S-частицу. Она включает
- малую субъединицу (40S), содержащую рибосомальную РНК с коэффициентом седиментации 18S рРНК и 30 молекул различных белков,
- большую субъединицу (60S), которая включает 3 разные молекулы рРНК (одну длинную и две короткие - 5S, 5,8S и 28S), а также 45 белковых молекул.
Субъединицы образуют "скелет" рибосомы, каждый из которых окружен своими белками. Коэффициент седиментации полной рибосомы не совпадает с суммой коэффициентов двух ее субъединиц, что связано с пространственной конфигурацией молекулы.
Устройство рибосом прокариотов и эукариотов примерно одинаковое. Отличаются они лишь молекулярной массой. Бактериальная рибосома имеет коэффициент cедиментации 70S и обозначается как 70S-частица, что указывает на меньшую скорость осаждения; содержит
- малую (30S) субъединицу - 16S рРНК + белки
- большую субъединицу (50S) - 23S рРНК + 5S рРНК + белки большой субчастицы (рис.)
В рРНК среди азотистых оснований выше чем обычно содержание гуанина и цитозина. Встречаются также минорные нуклеозиды, но не столь часто, как в тРНК: примерно 1 %. Это, в основном, нуклеозиды, метилированные по рибозе. Во вторичной структуре рРНК много двухцепочечных участков и петель (рис.). Таково строение молекул РНК, образуемых в двух последовательно проходящих процессах - транскрипции ДНК и созревании (процессинге) РНК.
Транскрипция рРНК с ДНК и процессинг рРНК
Пре-рРНК образуется в ядрышке, где находятся транскриптоны рРНК. Траснкрипция рРНК с ДНК происходит при помощи двух дополнительных РНК-полимераз. РНК-полимераза I транскрибирует 5S, 5,8S и 28S в виде одного длинного 45S-тpaнскрипта, который затем разделяется на необходимые части. Таким образом обеспечивается равное количество молекул. В организме человека в каждом гаплоидном геноме присутствует примерно 250 копий последовательности ДНК, кодирующей 45S-транскрипт. Они расположены в пяти кластерных тандемных повторах (т. е. попарно друг за другом) в коротких плечах хромосом 13, 14, 15, 21 и 22. Данные участки известны как ядрышковые организаторы, так как их транскрипция и последующий процессинг 45S-транскрипта происходят внутри ядрышка.
Не менее чем в трёх кластерах хромосомы 1 существует 2000 копий 5S-pPHK гена. Их транскрипция протекает в присутствии РНК-полимеразы III снаружи ядрышка.
В ходе процессинга остается чуть больше половины пре-рРНК и освобождаются зрелые рРНК. Часть нуклеотидов рРНК подвергается модификации, которая состоит в метилировании оснований. Реакция осуществляется метилтрансферазами. В роли донора метальных групп выступает S-аденозилметионин. Зрелые рРНК соединяются в ядре с белками рибосом, поступающих сюда из цитоплазмы, и образуют малую и большую субчастицы рибосом. Зрелые рРНК транспортируются из ядра в цитоплазму в комплексе с белком, который дополнительно защищает их от разрушения и способствует переносу.
Центры рибосом
Рибосомы существенно отличаются от других органел клетки. В цитоплазме они встречаются в двух состояниях: в неработающем, когда большая и малая субъединицы отделены друг от друга, и в активном - во время выполнения своей функции - синтеза протеина, когда субъединицы соединяются друг с другом.
Процесс соединения субъединиц рибосом или сборка активной рибосомы обозначается как инициация трансляции. Эта сборка происходит строго упорядоченным образом, что обеспечивается функциональными центрами рибосом. Все эти центры находятся на контактирующих поверхностях обеих субъединиц рибосомы. К ним относятся:
- Центр связывания мРНК (М центр). Он образован участком 18S рРНК, который комплементарен на протяжении 5-9 нуклеотидов 5"-нетранслируемому фрагменту мРНК
- Пептидильный центр (П-центр). В начале процесса трансляции с ним связывается инициирующая аа-тРНК. У эукариот инициирующий
кодон всех мРНК всегда кодирует метионин, поэтому инициирующей аа-тРНК является одна из двух метиониновых аа-тРНК, отмечаемая нижним индексом i:
Мет-тРНК i Met . На последующих же стадиях трансляции в П-центре находится пептидил-тРНК, содержащая уже синтезированную часть пептидной цепи.
Иногда говорят также о Е-центре (от "exit" - выход), куда перемещается тРНК, потерявшая связь с пептидилом, перед тем, как покинуть рибосому. Однако можно рассматривать этот центр как составную часть П-центра.
- Аминокислотный центр (А-центр) - место связывания очередной аа-тРНК.
- Пептидилтрансферазный центр (ПТФ центр) - он катализирует перенос пептидила из состава пептидил-тРНК на поступившую в А центр очередную аа-тРНК. При этом образуется еще одна пептидная связь и пептидил удлиняется на одну аминокислоту.
Как в аминокислотном центре, так и в пептидильном центре антикодоновая петля соответствующей тРНК (аа-тРНК или пептидил-тРНК), очевидно, обращена к М-центру - центру связывания матричной РНК (взаимодействуя с мРНК), а акцепторная петля с аминоацилом или пептидилом к ПТФ центру.
Распределение центров между субъединицами
Распределение центров между субъединицами рибосомы происходит следующим образом:
- Малая субъединица. Поскольку именно она содержит 18S-рРНК, с участком которой связывается мРНК, то М-центр расположен на данной субъединице. Кроме того, здесь же находятся основная часть А-центра и небольшая часть П-центра.
- Большая субъединица . На ее контактирующей поверхности расположены остальные части П- и A-центров. В случае П-центра - это его основная часть, а в случае А-центра - участок связывания акцепторной петли аа-тРНК с аминокислотным радикалом (аминоацилом); остальная же и большая часть аа-тРНК связывается с малой субъединицей. Большой субъединице принадлежит также ПТФ центр.
Инициация рибосомы (подготовка рибосомы к синтезу белка)
Синтез белка, или собственно трансляцию, принято разделять на три фазы: инициации (начало), элонгации (удлинение полипептидной цепи) и терминации (окончание). В фазу инициации происходит подготовка рибосомы к работе: соединение ее субъединиц. У бактериальных и эукариотических рибосом соединение субъединиц и начало трансляции протекает по-разному.
Начало трансляции - наиболее медленный процесс. В нем кроме субъединиц рибосомы, мРНК и тРНК принимают участие ГТФ и три белковых фактора инициации (IF-1, IF-2 и IF-3), которые не являются составными компонентами рибосомы. Факторы инициации облегчают связывание мРНК с малой субъединицей и ГТФ. ГТФ за счет гидролиза обеспечивает энергией процесс смыкания субъединиц рибосомы.

- Инициация начинается с того, что малая субъединица (40S) связывается с фактором инициации IF-3, в результате этого возникает препятствие к преждевременному связыванию большой субъединицы и возможность присоединения к ней мРНК.
- Далее к комплексу "малая субъединица (40S) + IF-3" присоединяется мРНК (своим 5"-нетранслируемым участком). При этом инициирующий кодон (АУГ) оказывается на уровне пептидильного центра будущей рибосомы.
- Далее к комплексу "малая субъединица + IF-3 + мРНК" присоединяются еще два фактора инициации: IF-1 и IF-2, при этом последний несет с собой особую транспортную
РНК, которую называют инициирующей аа-тРНК. В состав комплекса входит также ГТФ.
Малая субъединица соединяясь с мРНК представляет для считывания два кодона. На первом из них протеин IF-2 закрепляет инициаторную аа-тРНК. Второй кодон закрывает протеин IF-1, который блокирует его и не позволяет присоединиться следующей тРНК до момента полной сборки рибосомы.
- После связывания инициирующей аа-тРНК, т. е. Мет-тРНК i Met за счет комплементарного взаимодействия с мРНК (инициирующий кодон АУГ) и установки ее на свое место в П-центре происходит связывание субъединиц рибосомы. ГТФ гидролизуется до ГДФ и неорганического фосфата, а выдяляющаяся при разрыве данной макроэргической связи энергия создает термодинамический стимул для протекания процесса в нужном направлении. Одновременно факторы инициации покидают рибосому.
Таким образом, формируется своеобразный "бутерброд" из четырех основных компонентов. При этом в П-центре собранной рибосомы оказываются инициирующий кодон мРНК (АУГ) и связанная с ним инициирующая аа-тРНК. Последняя при образовании первой пептидной связи играет роль пептидил-тРНК.

Транскрипты РНК, синтезированные при помощи РНК-полимеразы, обычно претерпевают дальнейшие ферментативные превращения, называемые посттранскрипционным процессингом, и только после этого они обретают свою функциональную активность. Транскрипты незрелой матричной РНК носят название гетерогенной ядерной РНК (гяРНК). Они состоят из смеси очень длинных молекул РНК, содержащих интроны и экзоны. Созревание (процессинг) гяРНК у эукариотов включает несколько стадий, в одну из которых происходит удаление интронов - нетранслируемых вставочных последовательностей и сшивание экзонов. Процесс протекает таким образом, что следующие друг за другом экзоны, т. е. кодирующие фрагменты мРНК, никогда физически не разобщаются. Экзоны очень точно соединяются между собой с помощью молекул, называемых малыми ядерными РНК (мяРНК). Функция этих коротких ядерных РНК, состоящих приблизительно из ста нуклеотидов, долго оставалась непонятной. Ее удалось установить после того, как было обнаружено, что их нуклеотидная последовательность комплементарна последовательностям на концах каждого из интронов. В результате спаривания оснований, содержащихся в мяРНК и на концах свернутого в петлю интрона, последовательности двух экзонов сближаются таким образом, что становится возможным удаление разделяющего их интрона и ферментативное соединение (сплайсинг) кодирующих фрагментов (экзонов). Таким образом, молекулы мяРНК играют роль временных матриц, удерживающих близко друг от друга концы двух экзонов для того, чтобы сплайсинг произошел в правильном месте (рис.).
Превращение гяРНК в иРНК путём удаления интронов проходит в ядерном комплексе РНК-белков, называемом сплайсомой. У каждой сплайсомы есть ядро, состоящее из трёх малых (низкомолекулярных) ядерных рибонуклеопротеинов, или снурпов. Каждый снурп содержит хотя бы одну малую ядерную РНК и несколько белков. Существует несколько сотен различных малых ядерных РНК, транскрибируемых в основном РНК-полимеразой II. Считают, что их основная функция - распознавание специфических рибонуклеиновых последовательностей посредством спаривания оснований по типу РНК-РНК. Для процессинга гяРНК наиболее важны Ul, U2, U4/U6 и U5.
Митохондриальная РНК
Митохондриальная ДНК представляет собой непрерывную петлю и кодирует 13 полипептидов, 22 тРНК и 2 рРНК (16S и 23S). Большинство генов находятся на одной (тяжёлой) цепи, однако некоторое их количество расположено и на комплементарной ей лёгкой. При этом обе цепи транскрибируются в виде непрерывных транскриптов при помощи митохондриоспецифической РНК-полимеразы. Данный фермент кодируется ядерным геном. Длинные молекулы РНК затем расщепляются на 37 отдельных видов, а мРНК, рРНК и тРНК совместно транслируют 13 мРНК. Большое количество дополнительных белков, которые поступают в митохондрию из цитоплазмы, транслируются с ядерных генов. У пациентов с системной красной волчанкой обнаруживают антитела к снурп-белкам собственного организма. Кроме того, считают, что определённый набор генов малой ядерной РНК хромосомы 15q играет важную роль в патогенезе синдрома Прадера-Вилли (наследственное сочетание олигофрении, низкого роста, ожирения, гипотонии мышц).
Первичная структура РНК – порядок чередования рибонуклеозидмонофосфатов в полинуклеотидной цепи. В РНК, как и в ДНК, нуклеотиды связаны между собой 3",5"-фосфодиэфирными связями. Концы полинуклеотидных цепей РНК неодинаковы. На одном конце находится фосфорилированная ОН-группа 5"-углеродного атома, на другом конце – ОН-группа 3"-углеродного атома рибозы, поэтому концы называют 5"- и 3"-концами цепи РНК.
Вторичная структура РНК
Молекула рибонуклеиновой кислоты построена из одной полинуклеотидной цепи. Отдельные участки цепи РНК образуют спирализованные петли – «шпильки», за счёт водородных связей между комплементарными азотистыми основаниями A-U и G-C. Участки цепи РНК в таких спиральных структурах антипараллельны, но не всегда полностью комплементарны, в них встречаются неспаренные нуклеотидные остатки или даже одноцепочечные петли, не вписывающиеся в двойную спираль. Наличие спирализованных участков характерно для всех типов РНК.
Третичная структура РНК
Одноцепочечные РНК характеризуются компактной и упорядоченной третичной структурой, возникающей путём взаимодействия спирализованных элементов вторичной структуры. Так, возможно образование дополнительных водородных связей между нуклеотидными остатками, достаточно удалёнными друг от друга, или связей между ОН-группами остатков рибозы и основаниями. Третичная структура РНК стабилизирована ионами двухвалентных металлов, например ионами Mg 2+ , связывающимися не только с фосфатными группами, но и с основаниями.
Основные типы рнк
В цитоплазме клеток присутствуют 3 типа рибонуклеиновых кислот – транспортные РНК (тРНК), матричные РНК (мРНК) и рибосомальные РНК (рРНК). Они различаются по первичной структуре, молекулярной массе, конформации, продолжительности жизни и, самое главное, по функциональной активности.
http :// www . biochemistry . ru / biohimija _ severina / B 5873 Part 25-141. html
Методы определения первичной и вторичной структуры нуклеиновых кислот
Секвенирование – это общее название методов, которые позволяют установить последовательность нуклеотидов в молекуле ДНК. В настоящее время нет ни одного метода секвенирования, который бы работал для молекулы ДНК целиком; все они устроены так: сначала готовится большое число небольших участков ДНК (клонируется молекула ДНК многократно и «разрезается» её в случайных местах), а потом читается каждый участок по отдельности.
Клонирование происходит либо просто выращиванием клеток в чашке Петри, либо (в случаях, когда это было бы слишком медленно или по каким-то причинам не получилось бы) при помощи так называемой полимеразной цепной реакции. В кратком и неточном изложении работает она примерно так: сначала ДНК денатурируют, т.е. разрушают водородные связи, получая отдельные нити. Затем к ДНК присоединяют так называемые праймеры; это короткие участки ДНК, к которым может присоединиться ДНК-полимераза – соединение, которое, собственно, и занимается копированием (репликацией) нити ДНК . На следующем этапе полимераза копирует ДНК, после чего процесс можно повторять: после новой денатурации отдельных нитей будет уже вдвое больше, на третьем цикле – вчетверо, и так далее.
Все эти эффекты достигаются в основном с помощью изменений температуры смеси из ДНК, праймеров и полимеразы; для наших целей важно, что это достаточно точный процесс, и ошибки в нём редки, а на выходе получается большое число копий участков одной и той же ДНК. Разные методы секвенирования отличаются друг от друга не методами клонирования, а тем, как потом прочесть получившийся «суп» из многочисленных копий одной и той же ДНК.
Метод ДНК-ДНК гибридизации основан на том факте, что стабильность ДНК-ДНК дуплексов при определенной температуре зависит от числа нуклеотидов образующих комплементарные пары. Очевидно, что число комплементарных нуклеотидов в дуплексе где обе нити происходят из одной и той же молекулы ДНК (т.е. в гомодуплексах) равно 100%. Если же обе нити имеют разное происхождение (гетеродуплекс), то, в зависимости от числа произошедших мутаций, число комплементарных пар будет меньше 100%. Соответсвенно гетеродуплексы должны распадаться (плавится) при более низкой температуре, чем гомодуплексы. Причем, чем ниже температура плавления, тем больше различия в двух последовательностях. Температурная стабильность гибридной ДНК определяется температурой при которой 50% гибридной ДНК диссоциировалось в одноцепочечную форму. Затем эта температура сравнивается со средней температурой 50%-го плавления гомодуплексов обоих типов последовательностей участвующих в образовании гетеродуплекса, эта температура обычно обозначается Tm. Разница между медианной температурой плавления гетеро- и гомодуплексов обозначается как dTm. Показана линейная зависимость dTm от числа неспаренных оснований ( Britten et. al., 1974 ): p=cdTm. Константа c обычно определяется условиями проведения эксперимента и обычно варьирует от 0.01 до 0.015. Определение dTm требует большого числа повторений, т.к. велика экспериментальная ошибка.
Основным свойством ДНК является ее способность к репликации.
http :// postnauka . ru / longreads /468
1.9. Репликация ДНК, транскрипция, трансляция, обратная транскрипция. Амплификация ДНК. Биосинтез белка, аминокислотный код. Организация генов, строение генов у про- и эукариот, понятие о клонировании.
Репликация – это процесс самоудвоения молекул ДНК, происходящий под контролем ферментов. Репликация осуществляется перед каждым делением ядра. Начинается она с того, что спираль ДНК временно раскручивается под действием фермента ДНК-полимеразы. На каждой из цепей, образовавшихся после разрыва водородных связей, по принципу комплементарности синтезируется дочерняя цепь ДНК. Материалом для синтеза служат свободные нуклеотиды, которые есть в ядре.
Схема репликации ДНК
Таким образом, каждая полинуклеотидная цепь выполняет роль матрицы для новой комплементарной цепи (поэтому процесс удвоения молекул ДНК относится к реакциям матричного синтеза). В результате получается две молекулы ДНК, у каждой из которых одна цепь остается от родительской молекулы (половина), а другая – вновь синтезированная. Причем одна новая цепь синтезируются сплошной, а вторая – сначала в виде коротких фрагментов, которые затем сшиваются в длинную цепь специальным ферментом – ДНК-лигазой. В результате репликации две новые молекулы ДНК представляют собой точную копию исходной молекулы.
Биологический смысл репликации заключается в точной передаче наследственной информации от материнской клетки к дочерним, что и происходит при делении соматических клеток.
http :// sbio . info / page . php ? id =11
Литература:
1) Н. Грин, У. Стаут, Д. Тейлор – Биология.
2) З.А. Шабарова и А.А. богданов – Химия нуклеиновых кислот и их полимеров.
3) А.П. Пехов – Биология и общая гинетика.
4) А. Микельсон – Химия нуклеозидов и нуклеотидов.
5) З. Гауптман, Ю. Грефе, Х. Ремане – Органическая химия
Транскри́пция – это процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех живых клетках. Другими словами, это перенос генетической информации с ДНК на РНК.
Транскрипция катализируется ферментом ДНК-зависимой РНК-полимеразой. Процесс синтеза РНК протекает в направлении от 5"- к 3"- концу, то есть по матричной цепи ДНК РНК-полимераза движется в направлении 3"- 5".
Транскрипция состоит из стадий инициации, элонгации и терминации. Единицей транскрипции является транскриптон, фрагмент молекулы ДНК, состоящий из промотора, транскрибируемой части и терминатора.
Инициация транскрипции – это сложный процесс, зависящий от последовательности ДНК вблизи транскрибируемой последовательности (а у эукариот также и от более далеких участков генома - энхансеров и сайленсеров ) и от наличия или отсутствия различных белковых факторов .
Элонгация транскрипции
Момент перехода РНК-полимеразы от инициации транскрипции к элонгации точно не определен. Три основных биохимических события характеризуют этот переход в случае РНК-полимеразы кишечной палочки : отделение сигма-фактора, первая транслокация молекулы фермента вдоль матрицы и сильная стабилизация транскрипционного комплекса, который кроме РНК-полимеразы включает растущую цепь РНК и транскрибируемую ДНК. Эти же явления характерны и для РНК-полимераз эукариот. Переход от инициации к элонгации сопровождается разрывом связей между ферментом, промотором , факторами инициации транскрипции, а в ряде случаев – переходом РНК-полимеразы в состояние компетентности в отношении элонгации. Фаза элонгации заканчивается после освобождения растущего транскрипта и диссоциации фермента от матрицы (терминация).
На стадии элонгации в ДНК расплетено примерно 18 пар нуклеотидов . Примерно 12 нуклеотидов матричной нити ДНК образует гибридную спираль с растущим концом цепи РНК. По мере движения РНК-полимеразы по матрице впереди нее происходит расплетание, а позади – восстановление двойной спирали ДНК. Одновременно освобождается очередное звено растущей цепи РНК из комплекса с матрицей и РНК-полимеразой. Эти перемещения должны сопровождаться относительным вращением РНК-полимеразы и ДНК.