В состав бактериальной клетки входит. Строение бактерии: особенности
Клетка прокариотических организмов имеет сложное строго упорядоченное строение и обладает принципиальными особенностями ультраструктурной организации и химического состава.
Структурные компоненты бактериальной клетки делят на основные и временные (рис. 2). Основными структурами являются: клеточная стенка, цитоплазматическая мембрана с ее производными, цитоплазма с рибосомами и различными включениями, нуклеоид; временные — капсула, слизистый чехол, жгутики, ворсинки, эндоспоры, образующиеся лишь на определенных этапах жизненного цикла бактерий, у некоторых видов они отсутствуют полностью.
У прокариотической клетки структуры, расположенные снаружи от цитоплазматической мембраны, называют поверхностными (клеточная стенка, капсула, жгутики, ворсинки).
Термин «оболочка» в настоящее время используется для обозначения клеточной стенки и капсулы бактерий или только клеточной стенки, цитоплазматическая мембрана не входит в состав оболочки и относится к протопласту.
Клеточная стенка — важный структурный элемент бактериальной клетки, располагающийся между цитоплазматической мембраной и капсулой; у бескапсульных бактерий — это внешняя оболочка клетки. Она обязательна для всех прокариот, за исключением микоплазм и L-форм бактерий. Выполняет ряд функций: защищает бактерии от осмотического шока и других повреждающих факторов, определяет их форму, участвует в метаболизме; у многих видов патогенных бактерий токсична, содержит поверхностные антигены, а также несет на поверхности специфические рецепторы для фагов. В клеточной стенке бактерий имеются поры, которые участвуют в транспорте экзотоксинов и других экзобелков бактерий. Толщина клеточной стенки 10—100 нм, и на ее долю приходится от 5 до 50 % сухих веществ клетки.
Основным компонентом клеточной стенки бактерий является пептидогликан, или муреин (лат. murus — стенка), — опорный полимер, имеющий сетчатую структуру и образующий ригидный (жесткий) наружный каркас бактериальной клетки. Пептидогликан имеет основную цепь (остов), состоящую из чередующихся остатков N-ацстил-М-глюкозамина и N-ацетилмурамовой кислоты, соединенных 1,4-гликозидными связями, идентичные тетрапептидные боковые цепочки, прикрепляющиеся к молекулам N-ацстилмурамовой кислоты, и короткие поперечные пептидные мостики, связывающие полисахаридные цепи. Два типа связей (гликозидные и пептидные), которые соединяют субъединицы пептидогликана, придают этому гетерополимеру структуру молекулярной сети. Остов пептидогликанового слоя у всех видов бактерий одинаков; тетрапептидные белковые цепочки и пептидные (поперечные) у неодинаковых видов различны.
По тинкториальным свойствам все бактерии подразделяются на две группы: грамположительные и грамотрицателъные. В 1884 г. X. Грам предложил метод окраски, который был использован для дифференцирования бактерий. Сущность метода состоит в том, что грамположительные бактерии прочно фиксируют комплекс генцианвиолета и йода, не подвергаются обесцвечиванию этанолом и поэтому не воспринимают дополнительный краситель фуксин, оставаясь окрашенными в фиолетовый цвет. У грамотрицательных бактерий этот комплекс легко вымывается из клетки этанолом, и они при дополнительном нанесении фуксина окрашиваются в красный цвет. У некоторых бактерий положительная окраска по Граму наблюдается только в стадии активного роста. Способность прокариот окрашиваться по методу Грама или обесцвечиваться этанолом определяется спецификой химического состава и ультраструктуры их клеточной стенки. Пептидогликан у грамположительных бактерий — основной компонент клеточной стенки и составляет от 50 до 90 %, у грамотрицательных — 1 —10 %. Структурные микрофибриллы пептидогликана грамотрицательных бактерий сшиты менее компактно, поэтому поры в их пептидогликановом слое значительно шире, чем в молекулярном каркасе грамположитсльных бактерий. При такой структурной организации пептидогликана фиолетовый комплекс генцианвиолета и йода у грамотрицательных бактерий будет вымываться быстрее.
Клеточная стенка грамположительных бактерий плотно прилегает к цитоплазматической мембране, массивна, се толщина находится в пределах 20—100 нм. Для нее характерно наличие тейхоевых кислот, они связаны с пептидогликаном и представляют собой полимеры трехатомного спирта — глицерина или пятиатомного спирта — рибита, остатки которых соединены фосфодиэфирными связями. Тейхоевые кислоты связывают ионы магния и участвуют в транспорте их в клетку. В составе клеточной стенки грамположительных прокариот в небольших количествах также найдены полисахариды, белки и липиды.
Рис. 2. Схема строения прокариотической клетки:
1 — капсула; 2 — клеточная стенка; 3 — цитоплазматическая мембрана; 4 — нуклеоид; 5 — цитоплазма; 6 — хроматофоры; 7 — тилакоиды; 8 — мезосома; 9 — рибосомы; 10 — жгутики; 11— базальное тельце; 12 — пили; 13 — включение серы; 14 — капли жира; 15 — гранулы полифосфата; 16 — плазмида
Клеточная стенка грамотрицательных бактерий многослойна, толщина ее 14—17 нм. Внутренний слой — пептидогликан, который образует тонкую (2 нм) непрерывную сетку, окружающую клетку. Пептидогликан содержит только мезодиаминопимелиновую кислоту и не имеет лизина. Внешний слой клеточной стенки — наружная мембрана — состоит из фосфолипидов, липополисахарида, липопротеина и белков. В наружной мембране содержатся белки основы (матричные), они прочно связаны с пептидогликановым слоем. Одной из их функций является формирование в мембране гидрофильных пор, через которые осуществляется диффузия молекул с массой до 600, иногда 900. Матричные белки, кроме того, выполняют еще роль рецепторов для некоторых фагов. Липополисахарид (ЛПС) клеточных стенок грамотрицательных бактерий состоит из липида А и полисахарида. Токсичный для животных ЛПС получил название эндотоксина. Тейхоевые кислоты у грамотрицательных бактерий не обнаружены.
Структурные компоненты клеточной стенки грамотрицальных бактерий отграничены от цитоплазматической мембраны и разделены промежутком, называемым периплазмой или периплазматическим пространством.
Протопласты и сферопласты. Протопласты — формы прокариот, полностью лишенные клеточной стенки, образующиеся обычно у грамположительных бактерий. Сферопласты — бактерии с частично разрушенной клеточной стенкой. У них сохраняются элементы наружной мембраны. Наблюдаются у грамотрицательных бактерий и значительно реже у грамположительных. Образуются в результате разрушения пептидогликанового слоя литическими ферментами, например лизоцимом, или блокирования биосинтеза пептидогликана антибиотиком пенициллином и др. в среде с соответствующим осмотическим давлением.
Протопласты и сферопласты имеют сферическую или полусферическую форму и в 3—10 раз крупнее исходных клеток. В обычных условиях наступает осмотический лизис и они погибают. В условиях повышенного осмотического давления способны некоторое время переживать, расти и даже делиться. При снятии фактора, разрушающего пептидогликан, протопласты, как правило, отмирают, но могут превращаться в L-формы; сферопласты легко реверсируют в исходные бактерии, иногда трансформируются в L-формы или же гибнут.
L-Формы бактерий. Это фенотипические модификации, или мутанты, бактерий, частично или полностью утратившие способность синтезировать пептидогликан клеточной стенки. Таким образом, L-формы — бактерии, дефектные по клеточной стенке. Свое название они получили в связи с тем, что были выделены и описаны в институте Листера в Англии в 1935 г. Образуются при воздействии L-трансформирующих агентов — антибиотиков (пенициллина, полимиксина, бацитрацина, венкомицина, стрептомицина), аминокислот (глицина, метионина, лейцина и др.), фермента лизоцима, ультрафиолетовых и рентгеновых лучей. В отличие от протопластов и сферопластов L-формы обладают относительно высокой жизнеспособностью и выраженной способностью к репродукции. По морфологическим и культуральным свойствам они резко отличаются от исходных бактерий, что обусловлено утратой клеточной стенки и изменением метаболической активности.
L-Формы бактерий полиморфны. Встречаются элементарные тельца размером 0,2—1 мкм (минимальные репродуцирующие элементы), шары — 1—5, большие тела — 5—50, нити — до 4 мкм и более. Клетки L-форм имеют хорошо развитую систему внутрицитоплазматических мембран и миелиноподобные структуры. Вследствие дефекта клеточной стенки осмотически неустойчивы и их можно культивировать только на специальных средах с высоким осмотическим давлением; они проходят через бактериальные фильтры.
Различают стабильные и нестабильные L-формы бактерий. Первые полностью лишены ригидной клеточной стенки, что сближает их с протопластами; они крайне редко реверсируют в исходные бактериальные формы. Вторые могут обладать элементами клеточной стенки, в чем они проявляют сходство со сферопластами; в отсутствие фактора, вызвавшего их образование, реверсируют в исходные клетки.
Процесс образования L-форм получил название L-трансформации или L-индукции. Способностью к L-трансформации обладают практически все виды бактерий, в том числе и патогенные (возбудители бруцеллеза, туберкулеза, листерии и др.).
L-Формам придается большое значение в развитии хронических рецидивирующих инфекций, носительстве возбудителей, длительной персистенции их в организме. Доказана трансплацентарная инвазивность элементарных телец L-форм бактерий.
Инфекционный процесс, вызванный L-формами бактерий, характеризуется атипичностью, длительностью течения, тяжестью заболевания, трудно поддается химиотерапии.
Капсула — слизистый слой, расположенный над клеточной стенкой бактерии. Вещество капсулы четко отграничено от окружающей среды. В зависимости от толщины слоя и прочности соединения с бактериальной клеткой различают макрокапсулу, толщиной более 0,2 мкм, хорошо различимую в световом микроскопе, и микрокапсулу, толщиной менее 0,2 мкм, обнаруживаемую лишь при помощи электронного микроскопа или выявляемую химическими и иммунологическими методами. Макрокапсулу (истинную капсулу) образуют В. anlhracis, C1. perfringens, микрокапсулу — Escherichia coJi. Капсула не является обязательной структурой бактериальной клетки: потеря ее не приводит к гибели бактерии. Известны бескапсульные мутанты бактерий, например сибиреязвенный вакцинный штамм СТИ-1.
Вещество капсул состоит из высокогидрофильных мицелл, химический же состав их весьма разнообразен. Основные компоненты большинства капсул прокариот — гомо- или гетсрополисахариды (энтсробактерии и др.). У некоторых видов бацилл капсулы построены из полипептида. Так, в состав капсулы В. anthracis входит полипептид Д-глутаминовой кислоты (правовращающий изомер). В состав микрокапсулы микобактерий туберкулеза млекопитающих входят гликопептиды, представленные сложным эфиром трегалозы и миколовой кислоты (корд-фактор).
Синтез капсулы — сложный процесс и у различных прокариот имеет свои особенности; считают, что биополимеры капсулы синтезируются на наружной поверхности цитоплазматической мембраны и выделяются на поверхность клеточной стенки в определенных специфических ее участках.
Существуют бактерии, синтезирующие слизь, которая откладывается на поверхности клеточной стенки в виде бесструктурного слоя полисахаридной природы. Слизистое вещество, окружающее клетку, по толщине часто превосходит диаметр последней. У сапрофитной бактерии лейконостока наблюдается образование одной капсулы для многих особей. Такие скопления бактерий, заключенных в общую капсулу, называются зооглеями.
Капсула — полифункциональный органоид, выполняющий важную биологическую роль. Она является местом локализации капсульных антигенов, определяющих вирулентность, антигенную специфичность и иммуногенность бактерий. Утрата капсулы у патогенных бактерий резко снижает их вирулентность, например у бескапсульных штаммов бациллы антракса. Капсулы обеспечивают выживание бактерий, защищая их от механических повреждений, высыхания, заражения фагами, токсических веществ, а у патогенных форм — от действия защитных сил макроорганизма: инкапсулированные клетки плохо фагоцитируются. У некоторых видов бактерий, в том числе и патогенных, способствует прикреплению клеток к субстрату.
В ветеринарной микробиологии выявление капсулы используют в качестве дифференциального морфологического признака возбудителя при исследовании на сибирскую язву.
Для окрашивания капсул применяют специальные методы — Романовского — Гимзы, Гинса — Бурри, Ольта, Михина и др.
Микрокапсулу и слизистый слой определяют серологическими реакциями (РА), антигенные компоненты капсулы идентифицируют при помощи иммунофлюоресцентного метода (РИФ) и РДД.
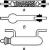
Жгутики — органоиды движения бактерий, представленные тонкими, длинными, нитевидными структурами белковой природы. Их длина превышает бактериальную клетку в несколько раз и составляет 10—20 мкм, а у некоторых спирилл достигает 80— 90 мкм. Нить жгутика (фибрилла) — полный спиральный цилиндр диаметром 12—20 нм. У вибрионов и протея нить окружена футляром толщиной 35 нм.
Жгутик состоит из трех частей: спиральной нити, крюка и базального тельца. Крюк — изогнутый белковый цилиндр, выполняющий функцию гибкого связывающего звена между базальным тельцем и жесткой нитью жгутика. Базальное тельце — сложная структура, состоящая из центрального стержня (оси) и колец.

Рис. 3. Жгутики:
а — монотрихи; б — амфитрихи; в — лофотрихи; г — перитрихи
Жгутики не являются жизненно важными структурами бактериальной клетки: существуют фазовые вариации бактерий, когда в одной фазе развития клетки они имеются, у другой — отсутствуют. Так, у возбудителя столбняка в старых культурах преобладают клетки без жгутиков.
Количество жгутиков (от I до 50 и более) и места их локализации у бактерий разных видов неодинаковы, но стабильны для одного вида. В зависимости от этого выделяют следующие группы жгутиковых бактерий: моиотрихи — бактерии с одним полярно расположенным жгутиком; амфитрихи — бактерии с двумя полярно расположенными жгутиками или имеющие по пучку жгутиков на обоих концах; лофотрихи — бактерии, имеющие пучок жгутиков на одном конце клетки; перитрихи — бактерии с множеством жгутиков, расположенных по бокам клетки или на всей ее поверхности (рис. 3). Бактерии, не имеющие жгутиков, называют атрихиями.
Будучи органами движения, жгутики типичны для плавающих палочковидных и извитых форм бактерий и лишь в единичных случаях встречаются у кокков. Они обеспечивают эффективное движение в жидкой среде и более медленное перемещение по поверхности твердых субстратов. Скорость движения монотрихов и лофотрихов достигает 50 мкм/с, амфитрихи и перитрихи движутся медленнее и обычно за 1 с проходят расстояние, равное размерам их клетки.
Бактерии передвигаются беспорядочно, однако они способны к направленным формам движения — таксисам, которые определяются внешними стимулами. Реагируя на различные факторы окружающей среды, бактерии за короткое время локализуются в оптимальной зоне обитания. Таксис может быть положительным и отрицательным. Принято различать: хемотаксис, аэротаксис, фототаксис, магнототаксис. Хемотаксис вызывается разницей в концентрации химических веществ в среде, аэротаксис — кислорода, фототаксис — интенсивностью освещения, магнитотаксис определяется способностью микроорганизмов ориентироваться в магнитном поле.
Выявление подвижных жгутиковых форм бактерий имеет значение для их идентификации при лабораторной диагностике инфекционных болезней.
Пили (фимбрии, ворсинки) — прямые, тонкие, полые белковые цилиндры толщиной 3—25 нм и длиной до 12 мкм, отходящие от поверхности бактериальной клетки. Образованы специфическим белком — пилином, берут начало от цитоплазматической мембраны, встречаются у подвижных и неподвижных форм бактерий и видимы только в электронном микроскопе (рис. 4). На поверхности клетки может быть от 1—2, 50—400 и более пилей до нескольких тысяч.

Рис. 4. Пили
Существует два класса пилей: половые (секспили) и пили общего типа, которые чаще называют фимбриями. У одной и той же бактерии могут быть пили разной природы. Половые пили возникают на поверхности бактерий в процессе конъюгации и выполняют функцию органелл, через которые происходит передача генетического материала (ДНК) от донора к реципиенту.
Пили общего типа располагаются перитрихиально (кишечная палочка) или на полюсах (псевдомонады); одна бактерия их может содержать сотни. Они принимают участие в слипании бактерий в агломераты, прикреплении микробов к различным субстратам, в том числе к клеткам (адгезивная функция), в транспорте метаболитов, а также способствуют образованию пленок на поверхности жидких сред; вызывают агглютинацию эритроцитов.
Цитоплазматическая мебрана и ее производные. Цитоплазматическая мембрана (плазмолемма) — полупроницаемая липопротеидная структура бактериальных клеток, отделяющая цитоплазму от клеточной стенки. Она является обязательным полифункциональным компонентом клетки и составляет 8—15 % ее сухой массы. Разрушение цитоплазматической мембраны приводит к гибели бактериальной клетки. На ультратонких срезах в электронном микроскопе выявляется ее трехслойное строение — два ограничивающих осмиофильных слоя, толщиной 2—3 нм каждый, и один осмиофобный центральный слой толщиной 4—5 нм.
Цитоплазматическая мембрана в химическом отношении — белково-липидный комплекс, состоящий из 50—75 % белков и 15—50 % липидов. Основная часть мембранных липидов (70— 90 %) представлена фосфолипидами. Она построена из двух мономолекулярных белковых слоев, между которыми расположен липидный слой, состоящий из двух рядов правильно ориентированных молекул липидов.
Цитоплазматичсская мембрана служит осмотическим барьером клетки, контролирует поступление питательных веществ в клетку и выход продуктов метаболизма наружу, в ней содержатся субстратспецифические ферменты-пермеазы, осуществляющие активный избирательный перенос органических и неорганических молекул.
Ферменты цитоплазматической мембраны катализуют конечные этапы синтеза мембранных липидов, компонентов клеточной стенки, капсулы и экзоферментов; на мембране локализованы ферменты окислительного фосфорилирования и ферменты транспорта электронов, ответственные за синтез энергии.
В процессе роста клетки цитоплазмзтическая мембрана образует многочисленные инвагинаты, формирующие внутрицитоплазмати-ческие мембраны структуры. Локальные инвагинаты мембраны получили название мезосом. Эти структуры хорошо выражены у грамположительных бактерий, хуже — у грамотрицательных и плохо — у риккетсий и микоплазм.
Установлена связь мезосом с хромосомой бактерии, такие структуры называются нуклеоидосомеши. Интегрированные с нуклеоидом мезосомы принимают участие в кариокинезе и цитокинезе микробных клеток, обеспечивая распределение генома после окончания репликации ДНК и последующее расхождение дочерних хромосом. Мезосомы, как и цитоплазматическая мембрана, являются центрами дыхательной активности бактерий, поэтому их иногда называют аналогами митохондрий. Однако значение мезосом окончательно еще не выяснено. Они увеличивают рабочую поверхность мембран, возможно, выполняют только структурную функцию, производя разделение бактериальной клетки на относительно обособленные отсеки, что создает более благоприятные условия для протекания ферментативных процессов. У патогенных бактерий обеспечивают транспорт белковых молекул экзотоксинов.
Цитоплазма — содержимое бактериальной клетки, отграниченное цитоплазматической мембраной. Состоит из цитозоля — гомогенной фракции, включающей растворимые компоненты РНК, вещества субстрата, ферменты, продукты метаболизма, и структурных элементов — рибосом, внутрицитоплазматических мембран, включений и нуклеоида.
Рибосомы — органоиды, осуществляющие биосинтез белка. Состоят из белка и РНК, соединенных в комплекс водородными и гидрофобными связями. Бактериальные рибосомы — гранулы диаметром 15—20 нм, имеют константу седиментации 70S и образованы из двух рибонуклеопротеидных субъединиц: 30S и 50S. Одна бактериальная клетка может содержать от 5000—50 000 рибосом, посредством и-РНК они объединяются в полисомы-агрегаты, состоящие из 50—55 рибосом, обладающих высокой белоксинтезирующей активностью.
В цитоплазме бактерий выявляются различного типа включения. Они могут быть твердыми, жидкими и газообразными, с белковой мембраной или без нее и присутствовать непостоянно. Значительная часть их представляет собой запасные питательные вещества и продукты клеточного метаболизма. К запасным питательным веществам относятся: полисахариды, липиды, полифосфаты, отложения серы и др. Из включений полисахаридной природы чаще обнаруживаются гликоген и крахмалоподобное вещество гранулеза, которые служат источником углерода и энергетическим материалом. Липиды накапливаются в клетках в виде гранул и капелек жира, к ним относятся окруженные мембраной гранулы поли-/3-оксимас-ляной кислоты, резко преломляющие свет и хорошо различимые в световом микроскопе. Выявляются и бациллы антракса и аэробных спорообразующих сапрофитных бактерий. Микобактерии в качестве запасных веществ накапливают воски. В клетках некоторых кори-небактерий, спирилл и других содержатся гранулы волютина, образованные полифосфатами. Они характеризуются метахромазией: толуидиновый синий и метиленовый синий окрашивают их в фиолетово-красный цвет. Волютиновые гранулы играют роль фосфатных депо.
К включениям, окруженным мембраной, также относятся газовые вакуоли, или аэросомы, они снижают удельную массу клеток, встречаются у водных прокариот.
Нуклеоид — ядро у прокариот. Он состоит из одной замкнутой в кольцо двухспиральной нити ДНК длиной 1,1 —1,6 нм, которую рассматривают как одиночную бактериальную хромосому, или генофор.
Нуклеоид у прокариот не отграничен от остальной части клетки мембраной — у него отсутствует ядерная оболочка.
В состав структур нуклеоида входят РНК-полимераза, основные белки и отсутствуют гистоны; хромосома закрепляется на цитоплазматической мембране, а у грамположительных бактерий — на мезосомс. Бактериальная хромосома реплицируется поликонсервативным способом: родительская двойная спираль ДНК раскручивается и на матрице каждой полинуклеотидной цепи собирается новая комплементарная цепочка. Нуклеоид не имеет митотического аппарата, и расхождение дочерних ядер обеспечивается ростом цитоплазматической мембраны.
Бактериальное ядро — дифференцированная структура. В зависимости от стадии развития клетки нуклеоид может быть дискретным (прерывистым) и состоять из отдельных фрагментов. Это связано с тем, что деление бактериальной клетки во времени осуществляется после завершения цикла репликации молекулы ДНК и оформления дочерних хромосом.
В нуклеоиде сосредоточен основной объем генетической информации бактериальной клетки.
Кроме нуклеоида в клетках многих бактерий обнаружены внехромосомные генетические элементы — плазмиды, представленные небольшими кольцевыми молекулами ДНК, способными к автономной репликации.
Мы даже не можем представить себе, сколько микроорганизмов постоянно окружают нас. Взявшись за поручень в автобусе, вы уже посадили себе на руку порядка ста тысяч бактерий, зайдя в общественный туалет, вы, опять-таки, наградили себя этими микроорганизмами. Бактерии всегда и везде сопровождают человека. Но не нужно на это слово реагировать негативно, ведь бактерии бывают не только патогенными, но также полезными для организма.
Ученые были очень удивлены, когда поняли, что некоторые бактерии сохранили свой внешний вид в течение приблизительно миллиарда лет. Такие микроорганизмы даже сравнивали с автомобилем марки "Фольксваген" - внешний вид одной их модели не менялся 40 лет, имея идеальную форму.
Бактерии появились на Земле одними из первых, поэтому их заслуженно можно назвать долгожителями. Интересным является тот факт, что эти клетки не имеют сформированного ядра, поэтому и по сей день привлекают много внимания к своему строению.
Что такое бактерии?
Бактерии - это микроскопические организмы растительного происхождения. Строение бактериальной клетки (таблица, схемы существуют для ясности понимания видов этих клеток) зависит от ее предназначения.
Эти клетки распространены везде, так как способны быстро размножаться. Существуют научные доказательства того, что буквально за шесть часов одна клетка может дать потомство в 250 тысяч бактерий. Эти одноклеточные организмы имеют множество разновидностей, которые различаются по форме.

Бактерии - очень живучие организмы, их споры могут сохранять способность к жизни на протяжении 30-40 лет. Переносятся эти споры с дуновением ветра, током воды и другими способами. Жизнеспособность сохраняется до температуры 100 градусов и при небольшом морозе. И все-таки, какое строение имеет бактериальная клетка? В таблице описаны основные составляющие бактерии, функции других органелл изложены ниже.
Шаровидные (кокки) бактерии
По своей природе они патогенные. Кокки делят на группы в зависимости от их расположения друг к другу:
- Микрококки (маленькие). Деление происходит в одной плоскости. Расположение в хаотичном одиночном порядке. Питаются готовыми органическими соединениями, но при этом не зависят от других организмов (сапрофиты).
- Диплококки (двойные). Делятся в такой же плоскости, что и микрококки, но образуют парные клетки. Внешне напоминают бобы или ланцетник.
- Стрептококки (в виде цепочки). Деление такое же, но клетки соединены между собой и выглядят, как бусы.
- Стафилококки (виноградная гроздь). Этот вид делится в нескольких плоскостях, при этом образуется скопление клеток, похожих на виноград.
- Тетракокки (четверка). Клетки делятся в двух перпендикулярных плоскостях, образуя тетрады.
- Сарцины (связка). Такие клетки делятся в трех плоскостях, которые взаимно перпендикулярны между собой. При этом внешне они похожи на пакеты или тюки, состоящие из множества особей четного количества.
Цилиндрические (палочки) бактерии
Палочки, которые образуют споры, подразделяют на клостридии и бациллы. По своим размерам эти бактерии бывают короткими и очень короткими. Конечные отделы палочек бывают закруглены, утолщены или обрезаны. В зависимости от расположения бактерий выделяют несколько групп: моно-, дипло- и стрептобактерии.
Спиралевидные (извитые) бактерии
Эти микроскопические клетки бывают двух видов:
- Вибрионы (с одиночным изгибом или вообще прямые).
- Спириллы (большие по размеру, но завитков мало).
Нитевидные бактерии. Существует две группы таких форм:
- Временные нити.
- Постоянные нити.
Особенности строения бактериальной клетки заключаются в том, что в процессе своего существования она способна изменять формы, но при этом полиморфизм не передается по наследству. Разные факторы действуют на клетку в процессе метаболизма в организме, вследствие этого наблюдаются количественные изменения в ее внешнем виде. Но как только действие извне прекратится, клетка примет прежний образ. Каковы особенности строения бактериальной клетки, можно выявить при ее рассмотрении с помощью микроскопа.
Строение бактериальной клетки, оболочка

Оболочка придает и поддерживает форму клетки, защищает внутренние составляющие от повреждений. Благодаря неполной проницаемости не все вещества могут попасть в клетку, что способствует обмену низко- и высокомолекулярных структур между внешней средой и самой клеткой. Также в стенке происходят различные химические реакции. С помощью электронного микроскопа нетрудно изучить, какое детализированное строение имеет бактериальная клетка.
Основа оболочки содержит полимер муреин. Грамположительные бактерии имеют однослойный скелет, состоящий из муреина. Здесь находятся полисахаридные и липопротеидные комплексы, фосфаты. У грамотрицатель-ных клеток муреиновый скелет имеет множество слоев. Наружный слой, прилегающий к клеточной стенке, является цитоплазматической мембра-ной. Она также имеет определенные слои, содержащие белки с липидами. Главная функция цитоплазматической мембраны - это контроль проникновения веществ внутрь клетки и выведения их (осмотический барьер). Это очень важная функция для клеток, так как с ее помощью происходит защита клеток.

Состав цитоплазмы
Живое полужидкое вещество, заполняющее клеточную полость, называется цитоплазмой. Большое количество белка, запас питательных веществ (жиры и жироподобные вещества) содержит в себе бактериальная клетка. Фото, сделанное во время исследования под микроскопом, хорошо показывает состав-ляющие части внутри цитоплазмы. В основной состав входят рибосомы, располагающиеся в хаотичном порядке и большом количестве. Также в составе имеются мезосомы, содержащие ферменты окислительно-вос-становительного характера. За счет них клетка черпает энергию. Ядро представлено в виде ядерного вещества, находящегося в тельцах хроматина.
Функции рибосом в клетках
Рибосомы состоят из субъединиц (2) и являются нуклеопротеидами. Соединяясь между собой, эти составляющие элементы образуют полисомы или полирибосомы. Главной задачей этих включений является белковый синтез, происходящий на основе генетической информации. Скорость седимента-ции 70S.
Особенности ядра бактерий

Генетический материал (ДНК) находится в неоформленном ядре (нуклеоид). Это ядро расположено в нескольких местах цитоплазмы, являясь неплотной оболочкой. Бактерии, име-ющие такое ядро, называются прокариотами. Аппарат ядра лишен мембраны, ядрышка и набора хромосом. А дезоксирибонуклеиновая кислота располагается в нем фибрильными пучками. Схема строения бактериальной клетки детально демонстрирует структуру ядерного аппарата.
При некоторых условиях у бактерий может возникнуть ослизнение оболочек. Вследствие этого проис-ходит образование капсулы. Если ослизнение очень сильное, то бактерии превращаются в зооглею (общая слизистая масса).

Капсула бактериальной клетки
Строение бактериальной клетки имеет особенность - это наличие защитной капсулы, состоящей из полисахаридов или гликопротеидов. Иногда эти капсулы состоят из полипептидов или клетчатки. Она располагается поверх клеточной оболочки. По толщине капсула может быть как толстой, так и тонкой. Ее образование происходит за счет условий, в которые попадает клетка. Основное свойство капсулы - это защита бактерии от высыхания.
Кроме защитной капсулы строение бактериальной клетки предусматривает ее двигательную ее способность.
Жгутики на бактериальных клетках

Жгутики являются дополнительными элементами, которые осуществляют движение клетки. Они представлены в виде нитей разной длины, которые состоят из флагеллина. Это белок, который обладает способностью сокращаться.
Состав жгутика трехкомпонентный (нить, крю-чок, базальное тельце). В зависимости от прикрепления и расположения выделили не-сколько групп подвижных бактерий:
- Монотрихи (эти клетки имеют 1 жгутик, расположенный полярно).
- Лофотрихи (жгутики в виде пучка на одном из концов клетки).
- Амфитрихи (пучки с обоих концов).
Существует много интересных фактов о бактериях. Так, уже давно доказано, что на мобильном телефоне содержится огромное количество этих клеток, даже на сидении унитаза их меньше. Другие бактерии позволяют нам качественно жить - питаться, выполнять определенную деятельность, без проблем освобождать свой организм от продуктов распада питательных веществ. Бактерии поистине разнообразны, их функции многогранны, но не следует забывать об их патологическом влиянии на организм, поэтому важно следить за собственной гигиеной и чистотой вокруг нас.
Общая схема строения бактериальной клетки показана на рисунке 2. Внутренняя организация бактериальной клетки сложна. Каждая систематическая группа микроорганизмов имеет свои специфические особенности строения.
Клеточная стенка. Клетка бактерий одета плотной оболочкой. Этот поверхностный слой, расположенный снаружи от цитоплазматической мембраны, называют клеточной стенкой (рис. 2, 14). Стенка выполняет защитную и опорную функции, а также придает клетке постоянную, характерную для нее форму (например, форму палочки или кокка) и представляет собой наружный скелет клетки. Эта плотная оболочка роднит бактерии с растительными клетками, что отличает их от животных клеток, имеющих мягкие оболочки. Внутри бактериальной клетки осмотическое давление в несколько раз, а иногда и в десятки раз выше, чем во внешней среде. Поэтому клетка быстро разорвалась бы, если бы она не была защищена такой плотной, жесткой структурой, как клеточная стенка.
Толщина клеточной стенки 0,01-0,04 мкм. Она составляет от 10 до 50% сухой массы бактерий. Количество материала, из которого построена клеточная стенка, изменяется в течение роста бактерий и обычно увеличивается с возрастом.
Основным структурным компонентом стенок, основой их жесткой структуры почти у всех исследованных до настоящего времени бактерий является муреин (гликопептид, мукопептид). Это органическое соединение сложного строения, в состав которого входят сахара, несущие азот,- аминосахара и 4-5 аминокислот. Причем аминокислоты клеточных стенок имеют необычную форму (D-стереоизомеры), которая в природе редко встречается.
 ,
,  ,
, 
Составные части клеточной стенки, ее компоненты, образуют сложную прочную структуру (рис. 3, 4 и 5).
С помощью способа окраски, впервые предложенного в 1884 г. Кристианом Грамом, бактерии могут быть разделены на две группы: грамположительные и грамотрицательные . Грамположительные организмы способны связывать некоторые анилиновые красители, такие, как кристаллический фиолетовый, и после обработки иодом, а затем спиртом (или ацетоном) сохранять комплекс иод-краситель. Те же бактерии, у которых под влиянием этилового спирта этот комплекс разрушается (клетки обесцвечиваются), относятся к грамотрицательным.
Химический состав клеточных стенок грамположительных и грамотрицательных бактерий различен.
У грамположительных бактерий в состав клеточных стенок входят, кроме мукопептидов, полисахариды (сложные, высокомолекулярные сахара), тейхоевые кислоты (сложные по составу и структуре соединения, состоящие из сахаров, спиртов, аминокислот и фосфорной кислоты). Полисахариды и тейхоевые кислоты связаны с каркасом стенок - муреином. Какую структуру образуют эти составные части клеточной стенки грамположительных бактерий, мы пока еще не знаем. С помощью электронных фотографий тонких срезов (слоистости) в стенках грамположительных бактерий не обнаружено. Вероятно, все эти вещества очень плотно связаны между собой.
Стенки грамотрицательных бактерий более сложные по химическому составу, в них содержится значительное количество липидов (жиров), связанных с белками и сахарами в сложные комплексы - липопротеиды и липополисахариды. Муреина в клеточных стенках грамотрицательных бактерий в целом меньше, чем у грамположительных бактерий. Структура стенки грамотрицательных бактерий также более сложная. С помощью электронного микроскопа было установлено, что стенки этих бактерий многослойные (рис. 6).

Внутренний слой состоит из муреина. Над ним находится более широкий слой из неплотно упакованных молекул белка. Этот слой в свою очередь покрыт слоем липополисахарида. Самый верхний слой состоит из липопротеидов.
Клеточная стенка проницаема: через нее питательные вещества свободно проходят в клетку, а продукты обмена выходят в окружающую среду. Крупные молекулы с большим молекулярным весом не проходят через оболочку.

Капсула. Клеточная стенка многих бактерий сверху окружена слоем слизистого материала - капсулой (рис. 7). Толщина капсулы может во много раз превосходить диаметр самой клетки, а иногда она настолько тонкая, что ее можно увидеть лишь через электронный микроскоп, - микрокапсула.
Капсула не является обязательной частью клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она служит защитным покровом клетки и участвует в водном обмене, предохраняя клетку от высыхания.
По химическому составу капсулы чаще всего представляют собой полисахариды. Иногда они состоят изгликопротеидов (сложные комплексы сахаров и белков) и полипептидов (род Bacillus), в редких случаях - из клетчатки (род Acetobacter).
Слизистые вещества, выделяемые в субстрат некоторыми бактериями, обусловливают, например, слизисто-тягучую консистенцию испорченного молока и пива.
Цитоплазма. Все содержимое клетки, за исключением ядра и клеточной стенки, называется цитоплазмой. В жидкой, бесструктурной фазе цитоплазмы (матриксе) находятся рибосомы, мембранные системы, митохондрии, пластиды и другие структуры, а также запасные питательные вещества. Цитоплазма обладает чрезвычайно сложной, тонкой структурой (слоистая, гранулярная). С помощью электронного микроскопа раскрыты многие интересные детали строения клетки.
, 
Внешний липопротвидный слой протопласта бактерий, обладающий особыми физическими и химическими свойствами, называется цитоплазматической мембраной (рис. 2, 15).
Внутри цитоплазмы находятся все жизненно важные структуры и органеллы.
Цитоплазматическая мембрана выполняет очень важную роль - регулирует поступление веществ в клетку и выделение наружу продуктов обмена.
Через мембрану питательные вещества могут поступать в клетку в результате активного биохимического процесса с участием ферментов. Кроме того, в мембране происходит синтез некоторых составных частей клетки, в основном компонентов клеточной стенки и капсулы. Наконец, в цитоплазматической мембране находятся важнейшие ферменты (биологические катализаторы). Упорядоченное расположение ферментов на мембранах позволяет регулировать их активность и предотвращать разрушение одних ферментов другими. С мембраной связаны рибосомы - структурные частицы, на которых синтезируется белок. Мембрана состоит из липопротеидов. Она достаточно прочна и может обеспечить временное существование клетки без оболочки. Цитоплазматическая мембрана составляет до 20% сухой массы клетки.
На электронных фотографиях тонких срезов бактерий цитоплазматическая мембрана представляется в виде непрерывного тяжа толщиной около 75A, состоящего из светлого слоя (липиды), заключенного между двумя более темными (белки). Каждый слой имеет ширину 20-30А. Такая мембрана называется элементарной (табл. 30, рис. 8).
 ,
, 
Между плазматической мембраной и клеточной стенкой имеется связь в виде десмозов - мостиков. Цитоплазматическая мембрана часто дает инвагинации - впячивания внутрь клетки. Эти впячивания образуют в цитоплазме особые мембранные структуры, названные мезосомами. Некоторые виды мезосом представляют собой тельца, отделенные от цитоплазмы собственной мембраной. Внутри таких мембранных мешочков упакованы многочисленные пузырьки и канальцы (рис. 2). Эти структуры выполняют у бактерий самые различные функции. Одни из этих структур - аналоги митохондрий. Другие выполняют функции зндоплазматической сети или аппарата Гольджи. Путем инвагинации цитоплазматической мембраны образуется также фотосинтезирующий аппарат бактерий. После впячивания цитоплазмы мембрана продолжает расти и образует стопки (табл. 30), которые по аналогии с гранулами хлоропластов растений называют стопками тилакоидов. В этих мембранах, часто заполняющих собой большую часть цитоплазмы бактериальной клетки, локализуются пигменты (бактериохлорофилл, каротиноиды) и ферменты (цитохромы), осуществляющие процесс фотосинтеза.
,
В цитоплазме бактерий содержатся рибосомы- белок-синтезирующие частицы диаметром 200А. В клетке их насчитывается больше тысячи. Состоят рибосомы из РНК и белка. У бактерий многие рибосомы расположены в цитоплазме свободно, некоторые из них могут быть связаны с мембранами.
Рибосомы являются центрами синтеза белка в клетке. При этом они часто соединяются между собой, образуя агрегаты, называемые полирибосомами или полисомами.
В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Однако их присутствие нельзя рассматривать как какой-то постоянный признак микроорганизма, обычно оно в значительной степени связано с физическими и химическими условиями среды. Многие цитоплазматические включения состоят из соединений, которые служат источником энергии и углерода. Эти запасные вещества образуются, когда организм снабжается достаточным количеством питательных веществ, и, наоборот, используются, когда организм попадает в условия, менее благоприятные в отношении питания.
У многих бактерий гранулы состоят из крахмала или других полисахаридов - гликогена и гранулезы. У некоторых бактерий при выращивании на богатой сахарами среде внутри клетки встречаются капельки жира. Другим широко распространенным типом гранулярных включений является волютин (метахроматиновые гранулы). Эти гранулы состоят из полиметафосфата (запасное вещество, включающее остатки фосфорной кислоты). Полиметафосфат служит источником фосфатных групп и энергии для организма. Бактерии чаще накапливают волютин в необычных условиях питания, например на среде, не содержащей серы. В цитоплазме некоторых серных бактерий находятся капельки серы.
Помимо различных структурных компонентов, цитоплазма состоит из жидкой части - растворимой фракции. В ней содержатся белки, различные ферменты, т-РНК, некоторые пигменты и низкомолекулярные соединения - сахара, аминокислоты.
В результате наличияв цитоплазме низкомолекулярных соединений возникает разность в осмотическом давлении клеточного содержимого и наружной среды, причем у разных микроорганизмов это давление может быть различным. Наибольшее осмотическое давление отмечено у грамположительных бактерий - 30 атм, у грамотрицательных бактерий оно гораздо ниже - 4-8 атм.
Ядерный аппарат. В центральной части клетки локализовано ядерное вещество - дезоксирибонуклеиновая кислот а (ДНК).
,
У бактерий нет такого ядра, как у высших организмов (эукариотов), а есть его аналог - «ядерный эквивалент» - нуклеоид (см. рис. 2, 8), который является эволюционно более примитивной формой организации ядерного вещества. Микроорганизмы, не имеющие настоящего ядра, а обладающие его аналогом, относятся к прокариотам. Все бактерии - прокариоты. В клетках большинства бактерий основное количество ДНК сконцентрировано в одном или нескольких местах. В клетках эукариотов ДНК находится в определенной структуре - ядре. Ядро окружено оболочкой- мембраной .
У бактерий ДНК упакована менее плотно, в отличие от истинных ядер; нуклеоид не обладает мембраной, ядрышком и набором хромосом. Бактериальная ДНК не связана с основными белками - гистонами - ив нуклеоиде расположена в виде пучка фибрилл.
Жгутики. На поверхности некоторых бактерий имеются придаточные структуры; наиболее широко распространенными из них являются жгутики - органы движения бактерий.
Жгутик закрепляется под цитоплазматической мембраной с помощью двух пар дисков. У бактерий может быть один, два или много жгутиков. Расположение их различно: на одном конце клетки, на двух, по всей поверхности и т. д. (рис. 9). Жгутики бактерий имеют диаметр 0,01-0,03 мкм, длина их может во много раз превосходить длину клетки. Бактериальные жгутики Состоят из белка - флагеллина - и представляют собой скрученные винтообразные нити.

На поверхности некоторых бактериальных клеток имеются тонкие ворсинки - фимбрии .
Жизнь растений: в 6-ти томах. - М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .
Форма. Различают несколько основных форм бактерий - кокковидные, палочковидные, извитые и ветвящиеся (рис.).
Шаровидные (кокковые) микробы по форме напоминают шар, но бывают овальные, плоские, односторонне вогнутые или слегка вытянутые. Шаровидные формы образуются в результате деления клеток в одной, двух, трех взаимно перпендикулярных или разных плоскостях. При делении клеток в одной плоскости клетки могут располагаться попарно, в связи с чем такие формы получили название диплококков. Если деление происходит последовательно в одной плоскости и клетки соединены в виде цепочки - это стрептококки (2). Деление кокка в двух взаимно перпендикулярных плоскостях ведет к образованию четырех клеток, или тетракокка. Пакетообразные кокки, или сарцины (3), - результат деления кокков в трех взаимно перпендикулярных плоскостях.
Палочковидные, или цилиндрические, формы принято делить на бактерии и бациллы (рисунок 3). Бактерии - палочковидные формы, не образующие спор (пишут Bact., например Bact. aceti). Бациллы - палочковидные формы, образующие споры (пишут Вас., например Вас. subtilis). Бактерии и бациллы бывают разными по форме и размерам. Концы палочек чаще закруглены, но могут быть срезаны под прямым углом (возбудитель сибирской язвы), иногда сужены.
Рисунок - Основные формы бактерий:
1- стафилококки; 2 - стрептококки; 3 - сарцины; 4 - гонококки; 5 - пневмококки; 6 - капсула пневмококков; 7 - коринебактерии дифтерии; 8 - клостридии; 9 – бациллы; 10 - вибрионы; 11 - спириллы; 12 - трепонемы; 13 - бореллии; 14 - лептоспиры; 15 - актиномицеты; 16 - расположение жгутиков: а - монотрих; б - лофотрих; в - амфитрих, г – перитрих
Среди палочковидных форм, образующих споры (бациллы), различают бациллы (9 ) и клостридии (8 ). Бациллы, за исключением Вас. anthracis , подвижны. Бациллы - аэробы. У бацилл споры не превышают толщины вегетативной клетки. Клостридии - анаэробы. Споры толще вегетативной клетки. Такие формы напоминают веретено, ракетку, лимон, барабанную палочку. Клостридии принимают участие во многих процессах в природе. Являются возбудителями анаэробных инфекций. Вызывают аммонификацию белковых веществ, мочевины. Разлагают фосфорорганические соединения. Фиксируют молекулярный азот и др.
Палочки, как и кокки, могут располагаться попарно или цепочкой. При соединении бактерий попарно образуются дипло-бактерии, при таком же соединении бацилл - диплобациллы. Соответственно образуются стрептобактерии и стрептобациллы, если клетки располагаются цепочкой. Тетрад и пакетов палочковидные формы не образуют, так как они делятся в одной плоскости, перпендикулярной продольной оси.
Извитые формы микробов определяют не только по длине и диаметру, но и по количеству завитков. Вибрионы(10) напоминают по форме запятую. Спириллы (11) - извитые формы, образующие до 5 завитков. Спирохеты - тонкие длинные извитые формы с множеством завитков. Они занимают промежуточное положение между бактериями и простейшими. Микобактерии - палочки с боковыми выростами (возбудители туберкулеза, паратуберкулеза). Коринебактерии напоминают микобактерии, но отличаются от них образующимися на концах утолщениями и включениями зерен в цитоплазме (дифтерийная палочка). Нитчатые бактерии - многоклеточные организмы, имеющие форму нити. Миксобактерии - скользящие микробы, по форме напоминающие палочки или веретено. Простекобактерии могут быть треугольной или иной формы. У некоторых из них лучевая симметрия. Свое название такие организмы получили по наличию остроконечных выростов - простек. Размножаются они делением, или почкованием.
Размеры. Размеры микроорганизмов определяются в микрометрах (мкм) (10 -6 м по системе СИ). Диаметр шаровидных форм 0,7-1,2 мкм; длина палочковидных 1,6-10 мкм, ширина 0,3-1 мкм. Вирусы - еще более мелкие существа. Их размеры определяются в нанометрах (1 нм = 10 -9 м). Нитчатые формы микробов достигают длины в несколько десятков микрометров. Для того чтобы представить размеры этих существ, достаточно сказать, что в одной капле воды может вмещаться несколько миллионов или миллиардов микроорганизмов.
Строение. Бактериальная клетка состоит из оболочки, наружный слой которой называется клеточная стенка, а внутренний - цитоплазматическая мембрана, а также цитоплазмы с включениями и нуклеоида. Имеются дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили, плазмиды; некоторые бактерии в неблагоприятных условиях способны образовывать споры.
Клеточная стенка - прочная, упругая структура, придающая бактерии определенную форму и вместе с подлежащей цитоплазматической мембраной «сдерживающая» высокое осмотическое давление в бактериальной клетке. Она защищает клетку от действии вредных факторов внешней среды, участвует в процессе ее деления и транспорте метаболитов.
Наиболее толстая клеточная стенка у грамположительных бактерий (до 50-60 нм); у грамотрицательных бактерий она составляет 15-20 нм.
В клеточной стенке грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков. Основным компонентом клеточной стенки грамположительных бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40-90% ее массы. У грамотрицательных бактерий количество пептидогликана в клеточной стенке - 5- 20%.
Цитоплазматическая мембрана прилегает к внутренней поверхности клеточной стенки бактерий и окружает наружную часть цитоплазмы. Она состоит из двойного слоя липидов, а также интегральных белков, пронизывающих ее насквозь. Цитоплазматическая мембрана участвует в регуляции осмотическогодавления, транспорте веществ и энергетическом метаболизмеклетки.
Цитоплазма бактериальной клетки представляет собой полужидкую, вязкую, коллоидную систему. Цитоплазма занимает основной объем бактериальной клетки и состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул - рибосом. В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, жирных кислот и полифосфатов (волютин).
Местами цитоплазма пронизана мембранными структурами - мезосомами , которые произошли от цитоплазматической мембраны и сохранили с ней связь. Мезосомы выполняют различные функции, в них и в связанной с ними цитоплазматической мембране расположены ферменты, участвующие в энергетических процессах - в снабжении клетки энергией.
Рибосомы рассеяны в цитоплазме в виде мелких гранул размером 20-30 нм; рибосомы состоят примерно наполовину из РНК и белка. Рибосомы ответственны за синтез белка клетки. В бактериальной клетке их может быть 5-50 тыс.
Нуклеоид - эквивалент ядра у бактерий. Он расположен в цитоплазмебактерий в виде двухнитчатой ДНК, замкнутой в кольцо и плотноуложенной наподобие клубка. В отличие от ядра эукариот нуклеоид бактерий не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Обычно в бактериальной клетке содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК.
Кроме нуклеоида в бактериальной клетке могут находиться внехромосомные факторы наследственности - плазмиды , представляющие собой ковалентно замкнутые кольца ДНК и способные к репликации независимо от бактериальной хромосомы.
Капсула - слизистая структура, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Обычно капсула состоит из полисахаридов, иногда из полипептидов, например, у сибиреязвенной бациллы. Капсула препятствует фагоцитозу бактерий. Капсулы присущи некоторым видам бактерий или могут образовываться при попадании микроба в макроорганизм.
Жгутики бактерий определяют подвижность клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, они прикреплены к цитоплазматической мембране и клеточной стенке специальными дисками, имеют большую длину, чем сама клетка. Они состоят из белка - флагеллина, закрученного в виде спирали.
Ворсинки, или пили (фимбрии) , - нитевидные образования, более тонкие и короткие, чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина. Они ответственны за прикрепление бактерий к поражаемой клетке, за питание, водно-солевой обмен; половые пили (F-пили) характерны для так называемых «мужских» клеток-доноров.
Споры - своеобразная форма покоящихся грамположительных бактерий, образующихся во внешней среде при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.). Процесс спорообразования проходит несколько стадий, в течение которых часть цитоплазмы и хромосома отделяются, окружаются цитоплазматической мембраной; образуется проспора, затем формируется многослойная, плохо проницаемая оболочка, придающая споре устойчивость к температуре и другим неблагоприятным факторам. При этом внутри одной бактерии образуется одна спора. Спорообразование способствует сохранению вида и не является способом размножения, как у грибов. Споры бактерий могут долго сохраняться в почве (возбудители сибирской язвы и столбняка - десятки лет). В благоприятных условиях споры прорастают, при этом из одной споры образуется одна бактерия.
Подвижность. Шаровидные бактерии, как правило, неподвижны. Палочковидные бактерии бывают как подвижными, так и неподвижными. Изогнутые и спиралевидные бактерии подвижны. Движение бактерий осуществляется с помощью жгутиков. Жгутики могут осуществлять вращательные движения. Наличие жгутиков, их расположение являются постоянным для вида признаком и имеют диагностическое значение. Скорость передвижения велика: за секунду клетка со жгутиками может пройти расстояние в 20-50 раз больше, чем длина ее тела.
Жгутики располагаются на поверхности тела бактерий по одиночке - монотрихиальное жгутикование , пучком на одном конце клетки - лофотрихиальное , пучком на обеих концах клетки - амфитрихиальное ; они могут находиться на всей поверхности клетки - перитрихиальное жгутикование . При неблагоприятных условиях жизни, при старении клетки, при механическом воздействии подвижность может быть утрачена.
Похожая информация.
Биологами XIX – начала ХХ века бактерии расценивались как примитивные организмы с точки зрения клеточной организации, считались крайним пределом жизни. Авторитетный немецкий ученый Кон писал, что бактерии «мельчайшие» и «простейшие» из всех живых форм образуют пограничную линию жизни, за пределами этих форм жизни не существует.
Однако, по мере развития науки была создана более совершенная микроскопическая техника и новые методы исследования. Применение современных методов исследований при изучении бактериальной клетки – электролитная и фазовоконтрастная микроскопия, дифференцированное центрифугирование, применение изотопов – позволили выявить отдельные клеточные структуры и выяснить их биологическую роль.
Бактериальная клетка имеет сложное строго упорядоченное строение. С анатомической точки зрения бактерия морфологически дифференцирована. В ней различают основные и временные структуры. К основным компонентам клетки относят клеточную стенку, цитоплазмотическую мембрану, цитоплазму с рибосомами, различные включения, нуклеоид. Эти структуры встречаются только на определенных стадиях развития бактерий.
Клеточная стенка – прочная, упругая структура, располагающаяся между цитоплазмотической мембраной и капсулой, а у бескапсульных видов бактерий – это внешняя оболочка клетки. клеточная стенка – тонкая бесцветная структура, она не видима в обыкновенный микроскоп без специальной обработки. Клеточная стенка придает бактериям постоянную форму и представляет собой скелет клетки. Ее можно рассматривать при световой микроскопии только у крупных форм бактерий. Например, у серобактерии Beggiatoa mirabilis стенка отчетливо видна и имеет двухконтурное строение. Стенку бактериальной клетки можно рассмотреть при плазмолизе в затемненном поле зрения микроскопа. Клеточной стенки не имеют микоплазмы и L-формы бактерий, для всех остальных прокариот она является обязательной структурой. Стенка клетки составляет в среднем 20 % сухого веса бактерий, толщина ее может достигать до 50 нм и более. Клеточная оболочка выполняет жизненно необходимые функции: защищает бактерию от повреждающих факторов внешней среды, осмотического шока, участвует в обмене веществ и в процессе деления клетки, содержит поверхностные антигены и специфические рецепторы для фагов, осуществляет транспортирование метаболитов. Оболочка бактерии полупроницаема, что обеспечивает избирательное проникновение питательных веществ в клетку из внешней среды. Опорный полимер клеточной стенки называется пептидогликаном (синонимы: мукопептид, муреин – от латинского murus – стенка) образует сетчатую структуру ковалентно связан тейховыми кислотами (от греческого teichos – стенка). При исследовании ультратонких срезов клеточной стенки было установлено, что она равномерно прилегает к подлежащим структурам, пронизана порами, благодаря которым осуществляется поступление различных веществ в клетку и, наоборот. Полученные фотограммы показали, что клеточная стенка характеризуется не одинаковой электроннооптической плотностью, т. е. обладает слоистостью. Стенка обрамляет бактерию, ее толщина и плотность одинаковы по всему периметру микробной клетки. На долю клеточной стенки приходится от 5 до 50 % сухих веществ клетки.
При изучении анатомии микроорганизмов с помощью светового микроскопа возникла необходимость в их окраске. Эта необходимость была реализована Х. Грамом, который в 1884 году предложил метод окраски, названный его именем и широко используемый для дифференцирования бактерий и в наше время. По отношению к окраске по Граму, все микроорганизмы подразделяются на две группы: грамположительные (грампозитивные) и грамотрицательные (грамнегативные). Сущность метода заключается в том, что грамположительные бактерии прочно связывают комплекс генцианвиолета и йода, который не обесцвечивается этанолом и не воспринимает дополнительный краситель фуксин, оставаясь окрашенными в сине-фиолетовый цвет. У грамотрицательных бактерий, упомянутый комплекс, вымывается из тела бактерий этанолом и они окрашиваются при обработке фуксином в красный цвет (цвет фуксина).
Такое окрашивание прокариот по Граму объясняется специфическим химическим составом и строением их клеточной стенки. Клеточная стенка грамположительных бактерий массивная, толстая (20-100 нм), плотно прилегает к цитоплазмотической мембране, большая часть ее химического состава представлена пептидогликаном (40-90 %), который связан с тейховыми кислотами. Стенка грамположительных микроорганизмов содержит в небольшом количестве полисахариды, липиды, белки. Структурные микрофибриллы пептидогликана сшиты прочно, компактно, поры в нем узкие и поэтому фиолетовый комплекс не вымывается, бактерии окрашиваются в сине-фиолетовый цвет.
Строение и состав грамотрицательных микроорганизмов характеризуется некоторыми особенностями. Клеточная стенка у грамнегативных бактерий тоньше, чем у грамположительных и составляет 14-17 нм. Она состоит из двух слоев: внешнего и внутреннего. Внутренний слой представлен пептидогликаном, который в виде тонкой (2 нм) непрерывной сетки опоясывает клетку. Пептидогликан у грамотрицательных бактерий составляет 1-10 %, микрофибриллы его сшиты менее прочно, чем у грамположительных бактерий, поры шире и поэтому комплекс генцианвиолета и йода вымывается из стенки этанолом, микроорганизмы окрашиваются в красный цвет (цвет дополнительного красителя – фуксина). Внешний слой содержит фосфолипиды, монополисахариды, липопротеин и белки. Липополисахарид (ЛПС) клеточных стенок грамотрицательных бактерий, токсичный для животных, получил название эндотоксина. Тейховые кислоты у грамотрицательных бактерий не обнаружены. Промежуток между клеточной стенкой и цитоплазмотической мембраной получил название периплазматического пространства, в котором содержатся ферменты.
Под влиянием лизоцима, пенициллина и других соединений синтез клеточной стенки нарушается и образуются клетки с измененной формой: протопласты – бактерии полностью лишенные клеточной стенки и сферопласты – бактерии с частично разрушенной клеточной стенкой. Протопласты и сферопласты имеют сферическую форму и в 3-10 раз крупнее исходных клеток. В условиях повышенного осмотического давления они могут расти и даже размножаться, а в обычных условиях наступает их лизис и гибель. При снятии ингибирующего фактора протопласты и сферопласты могут реверсировать в исходную форму, иногда трансформируются в L-формы бактерий. L-формы бактерий были выделены в 1935 году в институте Листера. Образуются они в результате воздействия на бактерии различного рода L-трансформирующих агентов (антибиотиков, аминокислот, ультрофиолетовых лучей, рентгенизлучения и т. д.). Это бактерии частично или полностью утратившие способность синтезировать пептидогликан клеточной стенки. По сравнению с протопластами и сферопластами они более устойчивы и обладают способностью к репродукции. Возбудители многих инфекционных болезней могут образовывать L-формы.
Цитоплазмотическая мембрана (плазмолемма) – полупроницаемая, трехслойная многопротеидная структура клетки, отграничивающая цитоплазму от клеточной стенки. Это обязательный компонент клетки, составляющий 8-15 % ее сухого вещества. При разрушении цитоплазмотической мембраны клетка гибнет. В химическом отношении мембрана представляет собой белково-липидный комплекс, состоящий из белков (50-70 %) и липидов (15-50 %). Цитоплазматическая мембрана выполняет важные функции в жизнедеятельности клетки. Она является осмотическим барьером клетки, участвует в процессах метаболизма, роста клетки, осуществляет избирательный перенос молекул органических и не органических веществ и т. д. В процессе роста клетки цитоплазматическая мембрана образует инвагинаты – выпячивания, которые получили название мезосом. Мезосомы хорошо выражены у грамположительных бактерий, хуже у грамотрицательных и совсем плохо у риккетсий и микоплазм. Мезосомы, связанные с нуклеоидом бактерии называются нуклеосомами. Они принимают участие в кариопинезе и кариокенезе микробной клетки. Значение мезосом окончательно не выяснено. Предполагают, что они принимают активное участие в процессе дыхания бактерий, поэтому их по аналогии сравнивают с митохондриями. Возможно, мезосомы выполняют структурную функцию и разделяя клетку на отдельные участки способствуют упорядоченности протекания обменных процессов.
Цитоплазма клетки представляет собой полужидкую массу, занимает основной объем бактерии, содержащий до 90 % воды. Состоит она из гомогенной фракции, называемой цитозолем, включающим структурные элементы – рибосомы, внутрицитоплазмотические мембраны, различного типа образования, нуклеоид. Кроме того в цитоплазме наличествуют растворимые компоненты РНК, вещества субстрата, ферменты, продукты метаболизма.
Цитоплазма образует внутреннюю среду клетки, которая объединяет все внутриклеточные структуры и обеспечивает взаимодействие их между собой.
Важнейший структурный компонент клетки проплазмотического типа – нуклеоид, который является аналогом ядра у эукариот. Он свободно располагается в цитоплазме, в центральной зоне клетки, представляет собой замкнутую в кольцо и плотно уложенную на подобие клубка двухнитчатую ДНК. Нуклеоид в отличае от четко оформленного ядра эукариот не имеет ядерной оболочки, ядрышков, основных белков (гистонов). Несмотря на это, считают, что нуклеоид – дифференцированная структура. В зависимости от функционального состояния клетки нуклеоид может быть дискретным и состоять из отдельных фрагментов. Дискретность его объясняется делением клетки и репликацией молекулы ДНК. ДНК нуклеоида является носителем генетической информации бактериальной клетки. При световой микроскопии нуклеоид может быть выявлен в результате окраски бактерий специальными методами (по Фельгену, по Романоскому-Гимзе). Кроме нуклеоида в клетках многих видов прокариот обнаружены внехромосомальные факторы наследственности – плазмиды, которые представляют собой молекулы ДНК, способные к автономной репликацией.
К органоидам клетки относят рибосомы – сферической формы рибонуклеиновые частицы диаметром 15-20 нм. Клетка прокариотического типа может содержать от 5 до 20 тысяч рибосом. Рибосома состоит из малой и большой субъединиц, обладающих по Свербергу константами сидиментации от 30 и 50 S, соответственно. Одна молекула матричной РНК обычно объединяет несколько рибосом наподобие бус, нанизанных на нить. Такие объединения рибосом называют полисомами. Рибосомы обладают высокой синтезирующей активностью, они синтезируют необходимые для жизнедеятельности микробной клетки белки.
В цитоплазме бактерий ваыявлены различного рода включения, которые бывают твердыми, жидкими и газообразными. Они представляют собой запасные питательные вещества (полисахариды, липиды, отложения серы и др.) и продукты обмена веществ.
Капсула – слизистая структура, толщиной более 0,2 мкм, связанная с клеточной стенкой и четко отграниченная от окружающей среды. Она выявляется при световой микроскопии в случае окрашивания бактерий специальными методами (по Ольту, Михину, Бурри-Гинсу). Многие бактерии образуют микрокапсулу – слизистое образование менее 0,2 мкм, выявленное только при электронной микроскопии или же химическими и иммунохимическими методами. Капсула не является обязательной структурой клетки, утрата ее не вызывает гибели бактерии. От капсулы необходимо отличать слизь – мукоидные экзополисахариды. Слизистые вещества откладываются на поверхности клетки, часто превосходя ее диаметр и не имеет четких границ.
Вещество капсул прокариот состоит в основном из гомо- или гетерополисахарид. У некоторых бактерий (например, лейконостока) в капсулу заключено несколько микробных клеток. Заключенные в одну капсулу бактерии представляют собой скопления называемые зоогелями.
Капсула выполняет важные биологические функции. В ней локализуются капсульные антигены, определяющие вирулентность, специфичность и иммуногенность бактерий. Капсула защищает микробную клетку от механических воздействий, высыхания, заражения фагами, токсических веществ, фагоцитоза. У некоторых видов бактерий, в том числе и патогенных, способствует прикреплению клеток к субстрату.
Жгутики являются органоидами движения бактерий. Они представляют собой тонкие, длинные, нитевидные структуры, состоящие из белка флагеллина (от латинского flagellum – жгутик). Этот белок обладает антигенной специфичностью. Длина жгутиков превышает длину бактериальной клетки в несколько раз и составляет 3-12 мкм, а толщина 12-20 нм. Жгутики прикреплены к цитоплазмотической мембране и клеточной стенке специальными дисками. Выявляют жгутики с помощью электронной микроскопии или же в световом микроскопе, но после обработки препаратов специальными методами. Жгутики не являются жизненноважными структурами клетки. Количество жгутиков различно у различных видов бактерий (от 1 до 50) и места их локализации тоже различны, но стабильны для каждого вида. В зависимости от локализации жгутиков различают: монотрихи – бактерии с одним полярно расположенным жгутиком; амфитирихи – бактерии с двумя полярно расположенными жгутиками, или по пучку жгутиков на каждом конце; лофотрихи – бактерии с пучком жгутиков на одном конце клетки; перитрихи – бактерии с множеством жгутиков, располагающихся по всему периметру клетки. Бактерии не имеющие жгутиков называются атрихиями. Жгутики типичны для плавающих палочковидных и извитых форм и в виде исключения встречаются у кокков. Монотрихи и лофотрихи движутся со скоростью 50 мкм в секунду. Обычно бактерии движутся беспорядочно. Под влиянием факторов внешней среды бактерии способны к направленным формам движения – таксисам. Таксис может быть положительным и отрицательным. Различают хемотаксис – обусловлен разницей концентрации химических веществ в среде, аэротаксис – кислорода, фототаксис – интенсивности освещения, магнитотаксис – характеризуется способностью микроорганизмов ориентироваться в магнитном поле.
Пили (ворсинки) – нитевидные образования более короткие, чем жгутики. Длина их достигает от 0,3 до 10 мкм, толщина 3-10 нм. Пили берут начало от цитоплазмотической мембраны, они обнаружены у подвижных и не подвижных форм микроорганизмов. Выявить их можно только с помощью электронной микроскопии. На поверхности бактериальной клетки может быть от 1-2 до нескольких десятков, сотен и даже тысяч пилей. Пили состоят из белка пилина, они обладают антигенной активностью.
Различают пили общего типа и половые. Первые ответственны за адгезию, т. е. прикрепление бактерий к поражаемой клетке, питание, водно-солевой обмен, слипание бактерий в агломераты, вторые – передачу наследственного материала (ДНК) от донора к реципиенту. У одного и того же вида бактерий могут быть пили обоих типов.
Споры (эндоспоры) – это особая форма покоящихся клеток, характеризующаяся резким снижением уровня метаболизма и высокой резистентностью. Споры образуются при неблагоприятных условиях существования бактерий. Внутри одной клетки образуется одна спора. Спорообразование наблюдается при дефиците питательных веществ, изменении рН, недостатке С, N, Р, высушивании, накоплении в окружающей клетку среде продуктов метаболизма и т. д. Споры характеризуются репрессией генома, анаболизмом, малым содержанием воды в цитоплазме, повышением концентрации катионов кальция, появление дипиколиновой кислоты.
Споры в поле зрения светового микроскопа имеют вид овальных, сильно преломляющих свет образований размером 0,8-1,5 мкм. Бактерии у которых размер споры не превышает диаметр клетки называются бациллами, а у которых превышает – клостридиями. Спора в клетке может располагаться центрально, ближе к концу – субтерминально, на конце бактерии – терминально. Строение споры сложное, но однотипное у разных видов бактерий. Центральная часть споры называется спороплазмой, в состав ее входят нуклеиновые кислоты, белки и дипиколиновая кислота. В спороплазме располагаются нуклеоид, рибосомы и нечетко выраженные мембранные структуры. Спороплазму обрамляет цитоплазмотическая мембрана, за которой следует зачаточный пептидогликановый слой, затем располагается массивный слой коры или иначе кортекса. На поверхности кортекса имеется наружная мембрана. Снаружи спора одета многослойной оболочкой, которая вместе со специфическими элементами споры и дипиколинатом кальция обуславливают ее устойчивость. Основное назначение спор – сохранение бактерий в неблагополучных условиях внешней среды. Споры устойчивы к действию высоких температур, химических веществ, могут длительное время существовать в покоящемся состоянии десятками и даже сотнями лет.
Видео: Ядро клетки листа комнатного растения undermicroscope